

Introduction
In the southeastern US production region of Alabama, Florida, and Georgia, weed control in peanut is accomplished with tactical measures of multiple herbicides with varying mechanism of action. This is accomplished via preemergence (PRE), at peanut emergence (AE), and postemergence (POST) herbicide applications. These timings are often dictated by the susceptibility of the weed species and crop safety.
Multiple applications of contact and residual herbicides are commonly used for peanut weed control as crop development and maturity require a long growing season (140-160 days). Diclosulam {N-(2,6-dichlorophenyl)-5-ethoxy-7-fluoro[1,2,4]triazolo[1,5-c]pyrimidine-2-sulfonamide} is an acetolactate synthase (ALS) inhibiting herbicide from the triazolopyrimidine sulfonanilide chemical family (Kleschick et al., 1992) registered since 2001 for peanut. Early research indicated that diclosulam provided effective weed control of dicot and nutsedge (Cyperus sp.) weed species in peanut when applied preplant incorporated (PPI), PRE, AE, or POST applications (Grey et al. 2001; Prostko et al., 1999; Bailey et al., 1998; Grichar et al., 1998; Prostko et al., 1998).
Historically, Benghal dayflower (Commelina benhgalensis L.) was not an issue in the southeastern US agronomic cropping systems. However, increased use of glyphosate in glyphosate-resistant crops from the mid-1990’s caused a weed shift in these production systems such that Benghal dayflower was prevalent by the early 2000’s (Webster et al. 2005; Webster and Nichols, 2012). Benghal dayflower is susceptible to several contact herbicides including paraquat, but this control is temporary in peanut due to lack of residual activity (Stephenson and Brecke, 2011). Benghal dayflower is also very fecund, producing seed from aerial and subterranean flowers that results in re-infestation, even after initial successful control in the same growing season (Sabila, et al. 2012; Webster et al. 2007).
University research noted that diclosulam, traditionally a PPI or PRE herbicide in peanut, could be used to control Benghal dayflower and other weeds when applied POST (Prostko et al., 2004; Lancaster et al. 2007; Everman et al. 2006; Grichar 2007). With this knowledge and support from Dow AgroSciences, University of Georgia researchers investigated and obtained a section 24(c) registration for Georgia, and was subsequently granted for Florida and Mississippi for peanut (Anonymous 2013). Research on diclosulam in peanut indicated variable control of many weed species in North Carolina; diclosulam POST application timing and rates experiments indicated variable control of common ragweed (Ambrosia artemisiifolia L.) from 60 to 100%, morningglory (Ipomoea sp. ) at 44 to 100%, and common lambsquarters (Chenopodium album L.) from 35 to 92% (Everman et al., 2006; Lancaster et al., 2007). Diclosulam applied in Georgia (Prostko et al., 2004) and Texas (Grichar 2007) controlled Florida beggarweed [Desmodium tortuosum (Sw.) DC.] less than 77%, and it did not control sicklepod [Senna obtusifolia (L.) Irwin & Barneby]. Palmer amaranth (Amaranthus palmeri S. Wats) control was variable (0 to 99%); this could be attributed to widespread ALS resistance in Georgia Palmer amaranth biotypes (Wise et al., 2009). However, extension recommendations from Georgia, Florida, and Alabama indicate that bristly starbur is extremely sensitive (>95% control) to diclosulam POST applications (Prostko, et al. 2004).
In 2009, diclosulam was applied PPI, PRE, or POST to over 100,000 ha of peanut in the southeast US (NAWQA, 2012). While diclosulam POST resulted in variable weed control of some species, bristly starbur control was unique and consistently near 100% regardless of size. The physiological factors that impart the extreme sensitivity of bristly starbur to diclosulam are not known.
Bristly starbur is a C3 plant native to tropical America, introduced to the US in the 1800’s and is now distributed in over 40 countries. The Latin binomial, Acanthospermum hispidum DC, is descriptive of the plant; Akanthos is Greek for spine, thorn, or prickle, sperma means seed, and hispidum is Latin for bristly or rough (Holm et al., 1997). The achene (also called a bur) is approximately 6 mm long, with stiff hooked spines, and two long spines at the apex (Figure 1). Seed are moved by clinging to equipment and animals, and both the vegetative parts of the plants and seed are poisonous to consume. In addition, the seed and plants are reported to contain allelochemicals (various phenolic acids) (Holm et al. 1997). Bristly starbur control is essential to prevent proliferation to other fields via contaminated harvesting equipment, peanut hay, and peanut seed given this species unique adapted ability to cling too many different surfaces. In 2009 and 2013 bristly starbur was the 6th and 10th most common and troublesome weed, respectively in Florida peanut and the 8th most common weed in Georgia peanut (Webster, 2009; 2013).

Bristly starbur (Acanthospermum hispidum) seed from Georgia (Photograph by Sidney Cromer, University of Georgia).
No reports of ALS-resistant bristly starbur have occurred. Target-site alteration as a mechanism of ALS-resistance in Palmer amaranth bitotypes from Georgia have been noted by substitutions at Trp574, Ser653, and Ala122 (Vencill et al., 2013). Weeds develop resistance to ALS-herbicides due to their widespread usage in multiple crops, the affinity for strong selection pressure, extreme weed sensitivity, and low use rates. Mutations in ALS conferring herbicide resistance are at least partially dominant and because the gene is nuclear inherited; it can be transmitted by seed and pollen (Tranel and Wright, 2002). Given that ALS-herbicide resistance is very common in many different weed species, there is concern that using diclosulam at reduced rates POST could negatively select for potential ALS-resistant bristly starbur.
While growers have now used diclosulam for several years to control weeds PPI, PRE, and POST in peanut, there are several questions as to why this herbicide is so effective on bristly starbur and document this physiology. Therefore, the objectives of this study were to: 1) evaluate bristly starbur dose response to diclosulam rates to establish the GR50 and I50 in greenhouse experiments, and 2) compare the absorption and mobility of 14C-diclosulam over time when applied POST to bristly starbur in laboratory experiments.
Materials and Methods
Multiple laboratory, greenhouse, and field experiments were conducted. Laboratory and greenhouse studies were conducted at the University of Georgia Weed Science Herbicide Research facilities located on the College of Agriculture Experiment Station in Tifton, GA (31.48° N, -83.529° W).
Seed Collection
Seed were collected from two geographically separate populations in Georgia in Terrell County (31.69 N, -81.52 W) and in Tattnall County (32.02 N, -82.22 W). These represented populations from the piedmont and coastal area of the state, respectively. Bristly starbur plants were collected based on a visual assessment of plant maturity. After greater than 50% leaf drop, entire plants were cut at the soil surface, placed into nylon bags, and then dried in a forced air chamber with no supplemental heat. After drying, plant stems were stripped of the seed heads by hand, threshed over sieves, and the remaining chaff removed with forced air. Cleaned seed were then placed into glass vials, labeled with the appropriate GPS coordinates, placed in controlled environmental storage, where they remained until testing.
Greenhouse and Laboratory Experiments
Laboratory testing of bristly starbur seed germination indicated approximately 80% for the Terrell County population and 50% for Tattnall County. After germination testing, bristly starbur seed were sown into 10 by 10 cm pots filled with a topsoil mix, watered daily, and fertilized (Miracle-Gro, P.O. Box 606, Marysville, OH 43040) weekly. After two weeks, pots were thinned to one plant per pot, which constituted an experimental unit. Greenhouse temperatures were regulated with diurnal settings of 32/25 (± 5 C) and included supplemental light provided by metal halide growth lights for 16 h per day.
Diclosulam Dose Response
In greenhouse studies, bristly starbur in the 4th - to 6th-leaf growth stage was treated with a POST application of six rates of diclosulam (0.2, 0.4, 0.8, 1.6, 3.3, and at 6.5g ai/ha) on a logarithmic dose using a common multiplier of 2 (Seefeldt et al., 1995). A non-treated control was also included in the treatment structure. For each diclosulam rate, foliar treatments were applied using deionized water and a compressed air sprayer calibrated to deliver 140 L/ha at 165 kPa. Nonionic surfactant (NIS) (80-20 Chem Nut Inc., Leesburg, GA 31763) at 0.25% (v/v) was included with all POST treatments. After treatment, all plants were returned to the greenhouse and maintained as previously described. Experiments were conducted three times; and all studies consisted of 4 to 8 replications per treatment. Each experiment was arranged as a randomized complete block design. Visual estimates of bristly starbur control were recorded 14 and 21 days after treatment (DAT), using a scale of 0 (no control) to 100 % (plant death). Above ground biomass was then harvested 21 DAT, dried in paper coin envelopes at 50 C for 72 hours and biomass measured.
Data were subjected to ANOVA and evaluated for experimental trial by population origin interactions. All data for dose response experiments were then subjected to ANOVA. Bristly starbur visual estimates of control and dry weight data were regressed against diclosulam rate (SAS 1999). The log-logistic regression equation was used to describe the dose response,

where C = lower limit, D = upper limit, b = slope, and I 50 = dose giving 50% response (for plant biomass growth response - GR50) (Seefeldt et al., 1995).
14C-diclosulam Translocation in Bristly Starbur
Plants were grown in the same manner as previously described. Two days prior to experiment initiation, bristly starbur plants were moved from the greenhouse into a growth chamber with a constant 28 C temperature and 50% relative humidity when they were in the 4th to 6th leaf stage of growth. Growth chamber lighting was provided by fluorescent and incandescent lamps at 450 μE/m2/s. The study was conducted as a randomized complete block design with treatments arranged as a split-plot and with 8 replicates. Whole plots were biotypes, and sub-plots were plant parts harvested. The study was conducted and then repeated twice. Since no differences were noted for the biotypes for the dose response experiments, plants from the two locations were combined for the 14C-diclosulam studies.
Prior to radiolabeling, the second fully expanded bristly starbur leaf (Li et al., 2005) was covered with polyethylene film. Then the entire plant was over sprayed with diclosulam at 0.8 g/ha mixed with deionized water and included a NIS at 0.25% (v/v), with a spray solution pH of 4.9. The film was then removed and the leaf was spotted with the radiolabeled solution using a microapplicator (Burkard Manufacturing Co. Ltd., Woodcock Hill Industrial Estate, Rickmansworth, Hertfordshire WD3 IPJ, U.K.). The spotting solution was prepared by mixing 0.5 ml of the spray solution with 14C-labeled diclosulam (100:20 v/v). Technical grade phenyl ring labeled-14C-diclosulam with 2.59 kBq/mg specific activity and 98% radiochemical purity was used. Ten 1-µl droplets of 14C-diclosulam were placed on the adaxial leaf surface approximately 2 mm away from the center vein, beginning at the base of the leaf and moving parallel to the center vein resulting in two lines of 5 drops on either side. Total specific activity applied contained 1.6 kBq of radioactivity. Plants were returned to the growth chamber immediately after spotting.
Bristly starbur plants were then harvested at 24 and 48 hour after treatment. Research on flumetsulam and other herbicides from the same triazolopyrimidine sulfonanilide chemical family as diclosulam, indicated maximum absorption occurred 16 to 30 hours after treatment (Frear et al., 1993; Hodges et al., 1990). Plants were cut at the soil line and sectioned into four parts: treated leaf, tissue above the treated leaf, tissue below the treated leaf, and roots. Soil was removed by washing the roots over a wire grid. Treated leaves were rinsed twice for 15 seconds with 5 ml of methanol:deionized water (1:1, v:v) to remove non-absorbed 14C-diclosulam (Li et al., 2005). A 1-ml aliquot of the each rinsate was added to 10 ml of scintillation fluid, and radioactivity was quantified by liquid scintillation spectrometry. All plant parts were dried for 48 hours at 45 C, weighed, and combusted with a biological sample oxidizer. Radioactivity in the oxidized samples was quantified by liquid scintillation spectrometry. The amount of herbicide absorbed was calculated as the total radioactivity recovered from oxidation of the four plant parts and expressed as a percent of the total radioactivity applied. Distribution of 14C-diclosulam in various plant parts was expressed as the percentage of total absorbed radioactivity.
All data for 14C-diclosulam experiments were subjected to ANOVA and tested for experiment interactions.
Results and Discussion
Dose Response
ANOVA indicated there was no treatment by bristly starbur population interactions; therefore data were combined across populations for regression using equation [1]. Diclosulam injury symptoms on bristly starbur included stunting, chlorosis, and necrosis 14 DAT, with some plant death by 21 DAT. The log-logistic dose response for diclosulam I50 for bristly starbur based on visual control ratings was 0.92 g a.i./ha (Figure 2). The diclosulam GR50 on a foliar tissue dry matter weight basis was 0.95 g a.i./ha (Figure 3). The registered field use rate for diclosulam POST is 27 g a.i./ha, indicating a susceptibility for bristly starbur of 29x and 28x for the I50 and GR50, respectively. Diclosulam molecular weight is 406.22 g/mole (Senseman, 2007), which corresponds to 457 µM at 26 g a.i./ha and 140 L/ha spray volume. Based on the application rates and spray solution volume for the dose response experiments, concentrations of diclosulam were 3.5, 7.1, 14.2, 29.6, 57.1, and 114.3 µM for the 0.2 0.4, 0.8, 1.6, 3.3, and 6.5 g a.i./ha, respectively. These data indicate the extreme sensitivity of bristly starbur to diclosulam as compared to other weeds that require the full use rate for peanut weed control including Benghal dayflower (Sabila et al., 2012), smallflower morningglory [Jacquemontia tamifolia (L.) Griseb.], common cocklebur (Xanthium strumarium L.), prickly sida (Sida spinosa L.) and common ragweed (Everman et al., 2006; Grey et al., 2001). Tolerant crop and weed species have limited translocation and rapid metabolism of diclosulam (Senseman, 2007; Hodges et al., 1990), which is similar to other sulfonamide herbicides (Frear et al., 1993). Data presented by Hodges et al. (1990) noted GR50 values of 0.8, 2.9, 2.7, and 2.1 g a.i./ha for redroot pigweed (Amaranthus retroflexus L.), velvetleaf (Abutilon theophrasti Medik.), jimsonweed (Datura stramonium L.), and tall morningglory (Ipomea purpurea (L.) Roth], respectively based on spray applications of 190 L/ha. These data are similar to the GR50 for bristly starbur in the current research of 0.33 g a.i./ha.
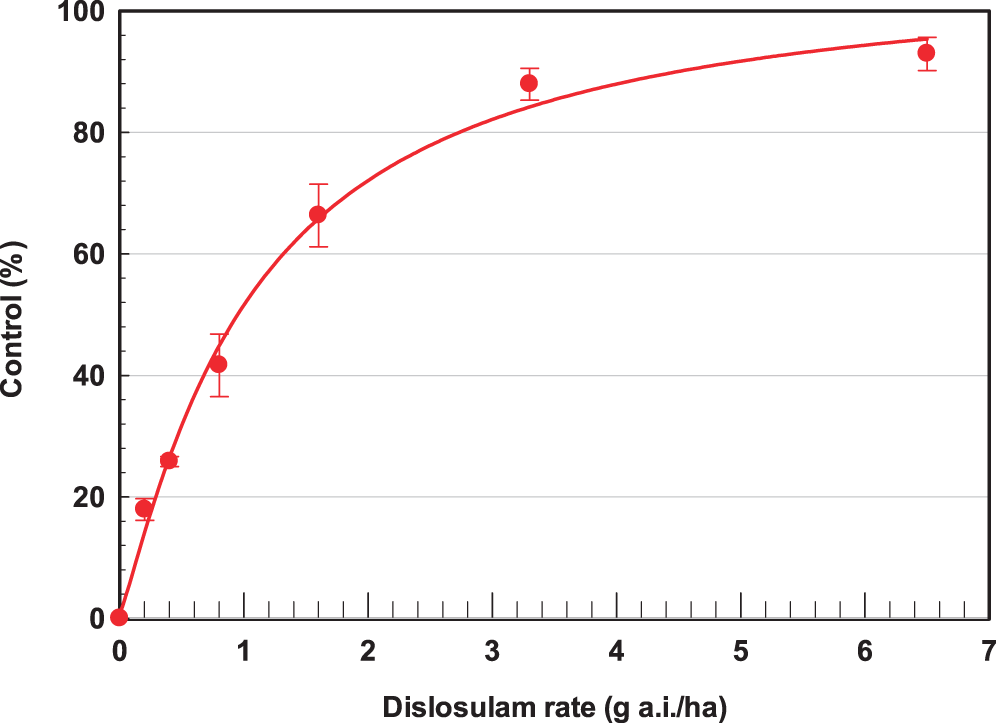
Log-logistic dose-response control curve and I50 value (number in parentheses represent the standard error) of bristly starbur (Acanthospermum hispidum) to diclosulam three weeks after application. The line represents the first-order regression equation and data points are the means of replications with bars indicating the standard error of the mean:
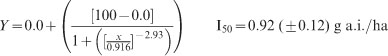
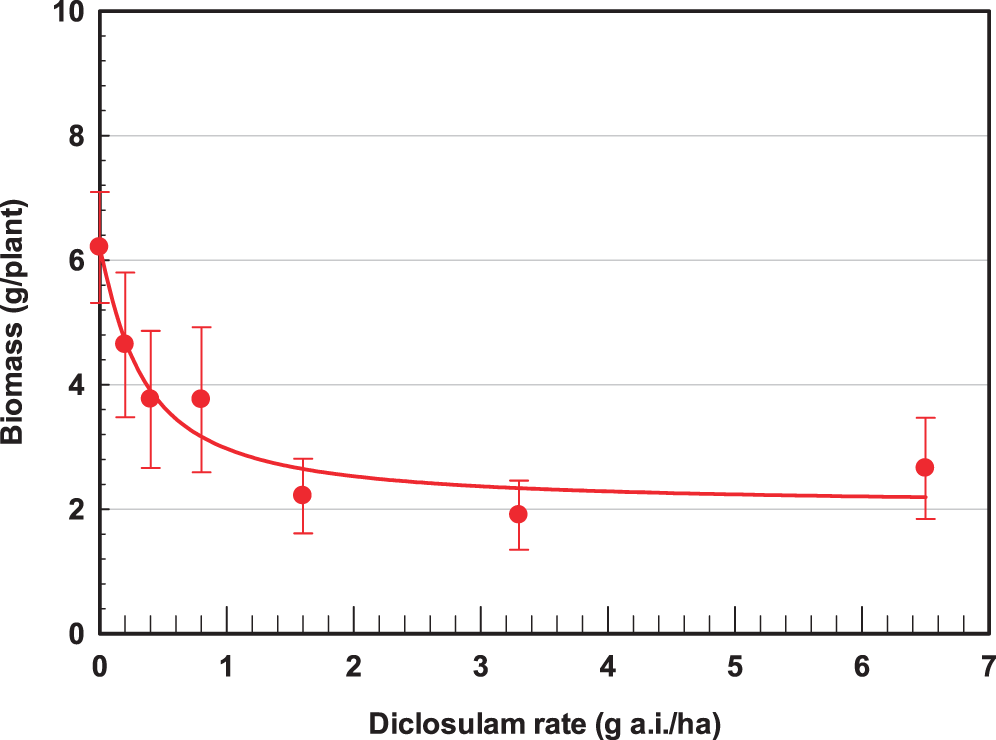
Log-logistic dose-response shoot dry weight curve and growth reduction GR50 value (number in parentheses represent the standard error) of bristly starbur (Acanthospermum hispidum) to diclosulam three weeks after application. The line represents the first-order regression equation and data points are the means of replications with bars indicating the standard error of the mean:
Based on these and previous data about the sensitivity of dicot species to diclosulam, it is suspected that bristly starbur does not readily metabolize diclosulam, resulting in low I50/GR50 values and extreme sensitivity (Figures 2 and 3). This level of diclosulam sensitivity in bristly starbur ((Prostko et al., 2004) can be an issue as ALS herbicide resistance is exacerbated by extremely sensitive weed species. Multiple Amaranthus species (McNaughton et al., 2005; Wise et al., 2009), common cocklebur (Schmidt et al., 2004), and common ragweed (Taylor et al., 2002) have been reported to have resistance to POST applied ALS herbicides. Additionally, weeds that have exhibited ALS resistance have point mutations within their genes encoding for ALS. Specifically, formally susceptible biotypes have Ala 122, Pro 197, Ala 205, Tip 574, or Ser 653 point mutations that are replaced by various other amino acids that have conferred ALS resistance (Tranel and Wright, 2002). In an effort to hinder the development of herbicide resistance, it will be pertinent in peanut to maintain current weed control PRE, AE, and POST herbicide applications utilizing multiple mechanisms of action to prevent selection for ALS resistance in bristly starbur.
14C-diclosulam Absorption and Translocation
There were no differences in plant biomass (mg per plant part and whole plant total) between the 24 and 48 hour after application timings (Table 1). Approximately 80% of the total applied radioactivity (1.28 of 1.6 kBq applied per plant) was recovered from leaf washes and oxidation of plant parts (n = 46 plants). Bristly starbur plants absorbed 65 and 55% of the applied 14C-diclosulam at 24 and 48 hours after treatment, respectively (Data not shown). There was no explanation as to why 10% less 14C-diclosulam was absorbed for the population of 48 hours plants verses the 24 hours treatments.
There were no differences in 14C-diclosulam as a percentage of absorption and translocation for the treated leaf, foliage above the treated leaf, foliage below the treated leaf, or the roots for the 24 and 48 hour after application harvest timings (Table 1). 14C-diclosulam moved acropetally out of the treated leaf 24 and 48 hr after treatment harvest (Table 1). Greater than 55% of the 14C-diclosulam absorbed translocated to the shoot above the treated leaf. Diclosulam is a weak acid (pKa = 4.09) (Senseman, 2007), and was anionic in charge for this experiment as the pH of the spray solution was 4.9. Upon application to the bristly starbur leaf surface, the anionic molecule was absorbed across the leaf cuticle via increased lipophilicity due to the halogens F and Cl on opposite ends of the diclosulam molecule (Jeschke, 2009). Once the diclosulam molecule entered the bristly starbur leaf cell, it was phloem translocated to the apical growing points. The F and Cl moieties increase the lipophicitiy and provide superior metabolic stability after phloem translocation that improves herbicidal activity (Jeschke 2009). This can partially explain the low diclosulam GR50 of 0.95 g a.i./ha for bristly starbur (Figure 3). When an herbicide has a Kow less than zero, they tend to be phloem translocated when foliar applied (Bromilow et al., 1990). The Kow for diclosulam at pH 7 is -0.047 (Senseman 2007), and this is a common plant intercellular pH. The molecular structure of diclosulam promotes lipophilic absorption across the leaf cuticle and intercellular pH of 7 and greater results in a negative Kow that increases phloem mobility. The data indicated that 14C-diclosulam had portioning ratios for above the treated leaf foliar tissue to roots of 65:1, and the above treated leaf foliar tissue to below treated leaf foliar tissue ratio of 6.6:1, with little to no root accumulation 24 hours after treatment. Similar results were observed with the 48 hour after treatment harvest.
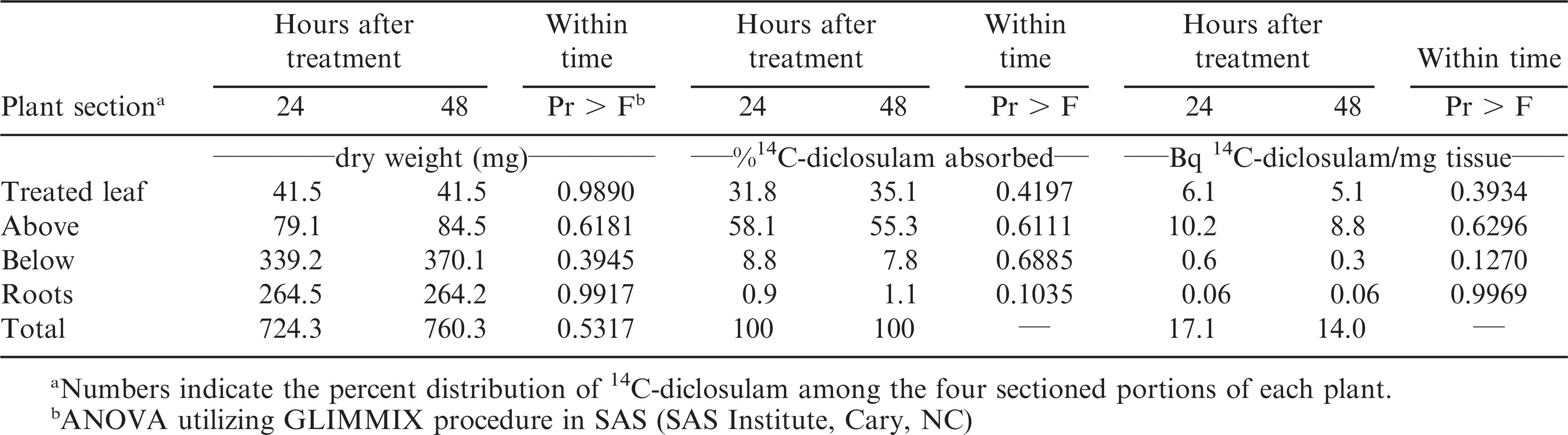
Plant biomass and distribution of 14C-diclosulam over time in bristly starbur (Acanthospermum hispidum) as a percentage of material absorbed and Becquerel per mg of plant tissue.
Conclusions
Bristly starbur exhibited extreme sensitivity to diclosulam and this efficacy was attributed to the lipophilic absorption and Kow phloem translocation of the molecule. While extremely effective for bristly starbur control, diclosulam is an ALS herbicide and weeds have previously demonstrated the ability to rapidly develop resistance to this mechanism of action due to point mutations. Therefore, it is highly recommended that when diclosulam is POST applied for Benghal dayflower control in peanut as registered, it is used in combination with other herbicides with different mechanisms of action that provide Benghal dayflower control. As most growers use PRE, AE, and POST herbicides in their production practices with multiple mechanisms of action, this should limit the potential for ALS resistant bristly starbur to occur. However, growers should always be aware of how weed populations change in their fields and respond appropriately to weed escapes as a means to minimize the occurrence, survival, and spread of herbicide resistant weeds.
Acknowledgements
The authors would like to thank Dow AgroSciences for the donation of radiolabeled 14C-diclosulam, and with the University of Georgia, Sidney Cromer for technical support, Jerry Davis for statistical analysis, and the staff at the Vidalia Onion and Vegetable Research Center for bristly starbur plant collection.
Literature Cited
Anonymous 2013 24(c) registration, GA0-40010 R-044-004 for suppression of tropical spiderwort in peanuts, Online at: http://www.cdms.net/LabelsMsds/LMDefault.aspx?pd=4332&t= Accessed Oct. 17, 2013 .
Bailey W.A Wilcut J.W Jordan D.L Askew S.D Hinton J.D and Langston V.B 1998 Evaluation of Strongram (DE 564, diclosulam) for weed control in southeastern peantus Proc. South. Weed Sci. Soc 51 : 59 (abstr).
Bromilow R.H Chamberlain K and Evans A.A 1990 Physicochemical aspects of phloem translocation of herbicides Weed Sci 38 : 305 – 314 .
Everman W.J Clewis S.B Taylor Z.G and Wilcut J.W 2006 Influence of diclosulam postemergence application timing on weed control and peanut tolerance Weed Technol 20 : 651 – 657 .
Frear D.S Swanson H.R and Tanaka F.S 1993 Metabolism of flumetsulam in wheat, corn, and barley Pest. Biochemistry and Phy 45 : 178 – 192 .
Grey T.L Bridges D.C and Eastin E.F 2001 Influence of application rate and timing of diclosulam on weed control in peanut Peanut Sci. 28 : 13 – 19 .
Grichar W.J 2007 Horse purslane, smellmelon, and Palmer amaranth control in peanut with postemergence herbicides Weed Technol 21 : 688 – 691 .
Grichar W.J Sestak D.C Brewer K.D Lemon R.L Dotray P.A and Hoelewyn T.A 1998 Nutsedge control in Texas peanut with diclosulam Proc. South. Weed Sci. Soc 51 : 58 – 59 (abstr).
Hodges C.C De Boer G.J , and Avalos J 1990 Uptake and metabolism as mechanisms of selective herbicidal activity of the 1,2,4-triazlo[1,5-a] pyrimidines Pesticide Sci 29 : 365 – 378 .
Holm L Doll J Holm E Pancho J and Herberger J 1997 World Weeds. Natural Histories and Distribution John Wiley & Sons, Inc. , 605 Third Avenue, New York, NY , 10158 .
Jeschke P 2009 The unique role of halogen substituents in the design of modern agrochemicals Pest Management Sci 66 : 10 – 27 .
Kleschick W.A Gerwick B.C Carson C.M Monte W.T and Snider S.W 1992 DE- 498, a new acetolactate synthase inhibiting herbicide with multicrop selectivity J. Agric. Food Chem 40 : 1083 – 1085 .
Lancaster S.H Bean J.B Lanier J.E Jordan D.L and Johnson P.D 2007 Compatibility ofdiclosulam with postemergence herbicides and fungicides Weed Technol 21 : 869 – 872 .
J Li Smeda R.J Sellers B.A and Johnson W.G 2005 Influene of formulation and glyphosate salt on absorption and translocation in three annual weeds Weed Scil 53 : 153 – 159 .
McNaughton K.E Letarte J Lee E.A and Tardif F.J 2005 Mutations in ALS confer herbicide resistance in redroot pigweed (Amaranthus retroflexus) and Powell amaranth (Amaranthus powellii) Weed Sci 53 : 17 – 22 .
National Water-Quality Assessment Program (NAWQA) 2012 Pesticide National Synthesis Project http://water.usgs.gov/nawqa/pnsp/. Accessed October 31, 2013.
Prostko E.P Grichar W.J Baughman T.A Brewer K.B and Lemon R.B 1999 Cadre and Strongarm comparisons for nutsedge (Cyperus spp.) control in peanuts-1998 Proc. Am. Peanut Educ. Res. Soc 31 : (abstr).
Prostko E.P Flanders J.T Komar S and Harrision E 2004 Strongarm applied postemergence in Georgia peanut Proc. Am. Peanut Educ. Res. Soc 36 : 30 (abstr).
Prostko E.P Lemon R.G Whitney R.A Grichar W.J Sestak D.C and Barrentine J.L 1998 Weed control in peanuts with diclosulam Proc. South. Weed Sci. Soc 51 : 59 (abstr).
Sabila M.H Grey T.L Webster T.M Vencill W.K and Shilling D.G 2012 Evaluation of factors that influence Benghal dayflower seed germination and emergence Weed Sci 60 : 75 – 80 .
[SAS] Statistical Analysis Systems 1999 SAS/STAT User's Guide, Version 8, Cary, NC Statistical Analysis System Institute Inc. , 3884 pp.
Schmidt L.A Talbert R.E and McClelland M 2004 Management of acetolactate synthase (ALS)–resistant common cocklebur in soybean Weed Technol 18 : 665 – 674 .
Seefeldt S.S Jensen J.E and Fruerst E.P 1995 Log-logistic analysis of herbicide doseresponse relationships Weed Technol 9 : 218 – 227 .
Senseman S.A 2007 Herbicide handboo Lawrence, KS : Weed Science Society of America . Pp. 62 – 64 .
Stephenson D.O IV and Brecke B.J 2011 Weed management in evenly- spaced 38- vs. 76- cm row peanut Peanut Sci 38 : 66 – 72 .
Taylor J.B Loux M.M Harrison S.K and Regnier E 2002 Response of ALS-Resistant common ragweed and giant ragweed to ALSinhibiting and alternative herbicides Weed Technol 16 : 815 – 825 .
Tranel P.J and Wright T.R 2002 Resistanc to wees to ALS-inhibiting herbicides What have we learned? Weed Sci 50 : 700 – 712 .
Vencill W.K Li X and Grey T.L 2013 Multiple mechanisms of Palmer amaranth resistance to ALS-inhibiting herbicides Proc. Weed Sci. Soc. America 57 : 363 (abstr.).
Webster TM 2009 Weed Survey-Southern States South. Weed Sci. Soc 62 : 515 – 517 .
Webster TM 2013 Weed Survey-Southern States South. Weed Sci. Soc 66 : 279 – 280 .
Webster T.M Burton M.G Culpepper A.S York A.C and Prostko E.P 2005 Tropical spiderwort: A tropical invader threatens agroecosystems of the Southern United States Weed Sci 53 : 501 – 508 .
Webster T.M Faircloth W.H Flanders J.T Prostko E.P and Grey T.L 2007 The critical period of Bengal dayflower control in peanut Weed Sci 55 : 359 – 364 .
Webster T.M and Nichols R.L 2012 Changes in the prevalence of weed species in the major agronomic crops of the Southern United States: 1994/1995 to 2008/2009 Weed Sc 60 : 145 – 157 .
Wise A Grey T.L Prostko E.P Vencill W.K and Webster T.M 2009 Establishing the geographical distribution and level of acetolactate synthase resistance of Palmer amaranth (Amaranthus palmeri) accessions in Georgia Weed Technol 23 : 214 – 220 .
Notes
- Professor and Professor, respectively. Department of Crop and Soil Sciences, The University of Georgia, 2360 Rainwater Drive, Tifton, GA 31793. *Corresponding author Email: tgrey@uga.edu.
Author Affiliations