

Introduction
Genomic science is an approach of investigating and understanding the highly complex structures and processes that make up a phenotype. The goal of genomics is to integrate knowledge about how the genome is organized, regulated, and interacted to create structures, products and activities. Genomic science takes advantage of large-scale and high throughput analyses to unravel the genetic make-up of an organism. Techniques to perform these tasks are continuing to be developed very rapidly, with the human genome project leading the way for other animal and plant systems. In plants, Arabidopsis has served as the model system, with rice (Oryza sativa L.), wheat (Triticum aestivum L. Thell.), maize (Zea mays L.), Medicago trancatula Gaertner, and a few other species leading the effort to develop methodologies and to sequence their genomes. Large data sets are compiled by the high thoughput instruments used to analyze genomes, and extensive bioinformatic analyses are required to sort, compare, and help interpret results. Unfortunately for the discipline, lists of analytical tools used in molecular biology and subsequent presentation of large data sets have tended to obscure fundamental significance of genomic science.
The research conducted in genomic science is highly labor intensive and expensive. Classical plant breeding programs, which are relatively inexpensive, are not well adapted for utilizing advanced technologies associated with genomics. Further, a large percentage of scientists who perform genomic research are mainly interested in the molecular function of specific genes or processes and are usually less interested in marker development for phenotypic selection applications. On the other hand, plant breeders need markers to facilitate selection and are generally not interested in developing large data sets for sequencing specific genes. Although the gap between the producer of genomic information (molecular biologist) and the user (plant breeders) is very wide, there is enormous potential for interactions among disciplines for plant improvement.
A fundamental question is “what can genomics deliver to improve agriculture productivity, quality, and safety”? Although efforts are needed to apply knowledge gained in molecular genetic research, data is generated to understand basic systems for genetic manipulation through transformation technologies, marker assisted selection, gene knockouts, and mutagenesis (Fig. 1). Genomics can identify relevant genetic pathways through transcriptional profiling and proteomics, thus leading to a better understanding of the plant functions that lead to phenotypic traits, plant tolerances, and pest resistances. Utilization of information from other plant species is possible by comparative genomic analysis, thus allowing the knowledge base of the majority of crops that are under-funded to be expanded with little cost. Lastly, high throughput techniques can be exploited to maximize research opportunities when a relatively small number of researchers are working on specific systems or problems. This paper will summarize the major events leading up to the current directions of genomic research in peanut and the current status of genomic-related science in the species.
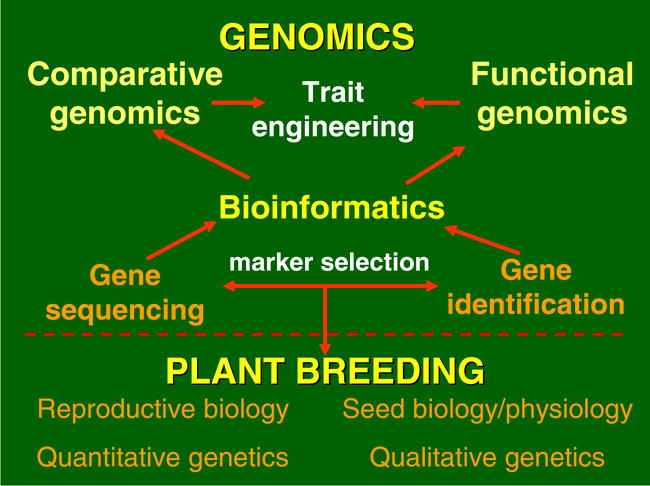
Background Information about Arachis hypogaea
The cultivated peanut, Arachis hypogaea L. is a polyploid species (2n = 2x = 40) with a large genome consisting of duplicate chromosomes. A hybridization event between two diploid species is believed to have occurred about 3500 years ago that led to the species we commonly know as peanut. Many species have been hypothesized as possible progenitors of A. hypogaea, and additional ones continue to be proposed from a variety of sources. However, the most likely ancestors are A. duranensis Krapov. and W. C. Gregory (A-genome) and A. ipaensis Krapov.and W. C. Gregory (B-genome) (Kochert et al., 1996). Arachis duranensis also is believed to be the female parent of the ancestral hybrid based on analyses of cytoplasmic genes (Hilu and Stalker, 1995). Temsch and Greihuber (2000) estimated that A. hypogaea has 5.91 pg DNA; and A. duranensis ranges between 2.49 and 2.87 pg DNA, depending on elevation and latitude of the accession (Temsch and Greihuber, 2001). Singh et al. (1996) concluded that the A and B genomes contributed nearly equal amounts of DNA to the domesticated peanut.
Although the genomes of several plant species are in the process of being sequenced, producing a complete DNA sequence-based map of peanut is unlikely because of its large genome size, the species being polyploid with duplicate genes in the two genomes, and the large resource base that will be required to complete the task (Paterson et al., 2004). The Arachis genome also has a large fraction of repetitive DNA, for example Dhillon et al. (1980) estimated that A. duranensis has 27% highly repetitive, 37% middle-repetitive, and 36% low-copy DNA. Alternatives to genome sequencing are available that can provide useful information about the plant genome, however, such as developing expressed sequence tag (EST) libraries. These libraries supply information about the genes being expressed (transcribed) in different tissues; although they do have limitations such as being a snapshot in time for gene activation, not providing information about gene regulation, and rare transcripts may not be detected. Importantly, the genes (regulatory and low abundance) may be important for many of the traits that distinguish peanut genotypes or separate it from other legumes (Paterson et al., 2004). Reviews have previously been published relating the history of molecular research in peanut (Stalker and Mozingo, 2001; Paterson et al., 2004) so only a brief account will be presented in this manuscript.
Marker Development in Peanut
Research with molecular aspects of the peanut genome began in the 1980s when protein and isozyme variation in A. hypogaea was determined to be of little use for characterizing variation within the cultivated peanut. Although large numbers of polymorphisms were detected among other species in the genus (Lu and Pickersgill, 1993; Stalker et al., 1994), the number of markers was too small to be routinely used in breeding programs. Restriction Fragment Length Polymorphisms (RFLPs) significantly expanded the number of data points available for analyses; but similar to isozymes and proteins, non-significant amounts of variation were found within the A. hypogaea gene pool (Kochert et al., 1991; Halward et al., 1991). Greater numbers of polymorphisms were found among Arachis species (Kochert et al., 1991; Paik-Ro et al., 1992) and were useful for clustering species of Arachis (Kochert et al., 1991). Further, intraspecific variation was found within the diploid A. duranensis (Stalker et al., 1995) and selected markers are useful for identifying individual accessions. The first molecular map of peanut was produced with RFLPs (Halward et al., 1993).
Development of Polymerase Chain Reaction (PCR) technologies, allowed DNA analyses with smaller aliquots of material and without the use of radioactivity. The commonly used Randomly Amplified Polymorphic DNA (RAPD) analysis for gene identification utilizes PCR technologies; but the method has the disadvantage of only detecting dominant markers. Observable differences by RAPD technologies within the cultivated peanut and among Arachis species are similar to reports for other marker systems (Halward et al., 1992). Amplified Fragment Length Polymorphisms (AFLPs) also are PCR-based technologies, but with greatly increased numbers of data points on electrophoretic gels. He and Prakash (1997) used 28 primer pairs to generate 111 AFLPs for DNA markers in A. hypogaea, with about 3% of the primers amplifying polymorphisms. AFLPs were the first molecular marker system used to differentiate closely related peanut cultivars (Herselman, 2003).
Hopkins et al. (1999) isolated the first Single Sequence Repeat (SSR) markers in peanut He et al. (2003) later reported that microsatellite markers developed from SSRs are more variable than other types of markers in peanut and 19 markers were polymorphic among A. hypogaea genotypes. Many hundreds of SSR makers have been developed (S. Knapp, personal commun.; Luo et al., 2005a), with about 30% being polymorphic among A. hypogaea lines (Ferguson et al., 2004; He et al., 2005; Moretzsohn et al., 2005). SSRs can be used to separate the cultivated lines (Moretzsohn et al., 2005) and they hold great potential for developing useful marker systems for improving selection efficiency in peanut. To date, the number of genes associated with molecular markers in peanut is small (Table 1), but the large number of molecular markers becoming available have great potential for utilizing in a crop improvement program. The next step is for plant breeders to develop appropriate plant populations for testing markers to establish associations with traits of interest. This will require cooperation between breeding and molecular genetic programs. These technologies have the greatest potential for traits that are complex, multigenic, and/or that are difficult to select in the greenhouse or field.
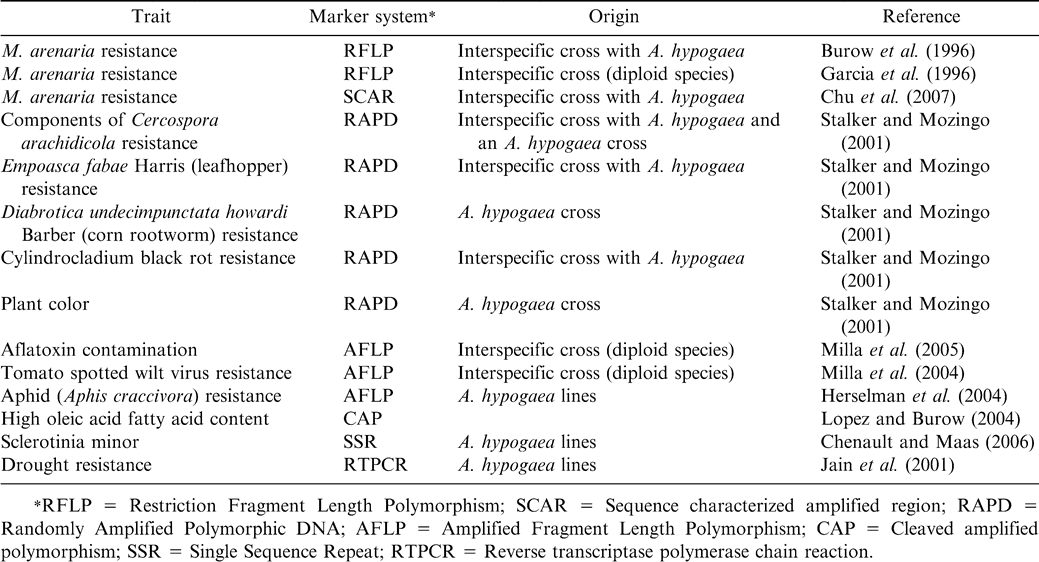
Genes associated with molecular markers in peanut.
In addition to markers being useful for associating with specific traits, they also may be useful for following introgression from Arachis species to A. hypogaea. This is important because recombination between the cultivated genomes and those of other species is rare, thus restricting selection for desired traits in interspecific hybrid derivatives (Holbrook and Stalker, 2003). Introgression is possible, however, because Garcia et al. (1995) observed genes from the A-genome species A. cardenasii Krapov. and W. C. Gregory in advanced generation A. hypogaea hybrids in 10 of 11 linkage groups on the diploid RFLP map developed by Halward et al. (1993).
Genomic Affinities and Speciation
Studies to understand the phylogenetic relationships among species in section Arachis, and to a lesser extent in other groups, have been made using isozyme variation (Stalker et al., 1994; Lu and Pickersgill, 1993), seed storage proteins (Singh et al., 1991; Bianchi-Hall et al., 1993; Liang et al. 2006), RFLPs (Kochert et al., 1991; Halward et al., 1991; Paik-Ro et al., 1992), SSRs (Hopkins et al., 1999), RAPDS (Halward et al., 1992; Lanham et al., 1992; Hilu and Stalker, 1995), and in situ hybridization (Raina and Mukai, 1999). The accumulated molecular data supports the hypothesis that A. hypogaea originated from a single hybridization event followed by chromosome doubling, and since its origin there has been very little to no introgression from diploid species into the cultivated peanut. Differences exist among marker systems when associating individual accessions within a species and for determining genetic distance between species; but in general, species cluster into genomic groups, with the B and D genomes being more related to each other than to the A genome. Polyploidy has evolved independently in the section Arachis (i.e., A. hypogaea and A. monticola Krapov. and Rigoni) and in species of section Rhizomatosae (Smartt and Stalker, 1982); and Nelson et al. (2006) used RAPD methodology to conclude that the Rhizomatosae species A. glabrata Benth. and A. pseudovillosa (Chodat and Hassl.) Krapov. and W. C. Gregory also may have evolved independently.
Molecular Maps of Peanut
Several molecular maps have been produced in peanut with different marker systems. The first map used RFLPs and utilized variation between the diploid species A. stenosperma Krapov. and W. C. Gregory × A. cardenasii where a total of 117 RFLP markers were mapped into 11 linkage groups (Halward et al., 1993). Garcia et al. (2005) developed a RAPD-based linkage map of peanut based on a backcross population [A. stenosperma × (A. stenosperma × A. cardenasii)] where 167 RAPD loci and 39 RFLPs were mapped to 11 linkage groups and all common markers mapped to the same linkage groups and mostly in the same order as in the map previously developed by Halward et al. (1993).
Another RFLP map was created by using progenies of a cross between A. hypogaea and TxAG-6 {a polyploidy of one B-genome and two A-genome diploid species: 4x[A. batizocoi Krapov. and W. C. Gregory × (A. cardenasii × A. diogoi Hoehne)]} where 383 markers were mapped (Burow et al., 1996). Genetic inheritance was disomic (with the exception of 1 linkage group), and the marker R239 was associated with nematode resistance. This marker mapped to the same linkage group in the map produced by Halward et al. (1993). Although the Halward et al. (1993), Burow et al. (1996), and Garcia et al. (2005) maps represent a good starting point for unraveling the peanut genome, they do not have sufficient numbers of markers to be highly useful for genetic studies and, because they are based on diploid species, there are problems relating the information to the cultivated peanut.
A microsatellite map with markers grouped into 11 linkages was made by Moretzsohn et al. (2005) by utilizing a cross between A. duranensis and A. stenosperma (both A-genome species). A second map with the B-genome species A. ipaensis and A. magna Krapov., W. C. Gregory, and C. E. Simpson used the same markers and a comparison between the A- and B-genome maps indicated that they were generally collinear (Moretzsohn et al., 2005). Lastly, Herselman et al. (2004) mapped 12 AFLP markers into five linkage groups by using A. hypogaea crosses.
Creating a comprehensive physical map of A. hypogaea will require a very large-scale effort (Paterson et al., 2004) and will require a library of large-insert DNA clones. Yüksel and Paterson (2005) produced a bacterial artificial clone (BAC) library with about 180,000 clones using the tetraploid peanut cultivar ‘Florunner’. The BAC library should serve as a highly useful resource for developing a physical map, but a limitation is the presence of duplicated copies of genes from the two progenitor species and Paterson et al. (2004) concluded that it will be difficult to distinguish true homologes vs. homoeologs from related species in BACs by hybridization-based molecular approaches. Although Lin et al. (2000) developed a method to determine the subgenome-specificity of individual BAC clones in polyploid species libraries, an alternative may be to produce BAC libraries for diploid progenitors of peanut and then compare them with the A. hypogaea libraries (Paterson et al., 2004).
TILLING Research in Peanut
Targeting Induced Local Lesions IN Genomes (TILLING) is a method developed to find genes of interest in a mutant population of a species. Because allergens in peanut cause a significant health problem in the human population, a high priority of the peanut industry is to eliminate or suppress the proteins that cause allergens. There are multiple seed storage proteins that give rise to allergens, with the major ones being Ara h 1, Ara h 2, and Ara h 3 (Burks et al., 1998); Ara h 2 is the most important peanut allergen (Koppelman et al., 2004). Ozias-Akins (pers. commun.) is developing a TILLING population in peanut with the goal of eliminating Ara h 2 and possibly other allergen genes.
Peanut Transformation
Ozias-Akins et al. (1993) reported the first successful transformation of peanut, with accompanying plant regeneration, by utilizing the micro-bombardment technique. Micro-bombardment has since been completed in peanut with a number of genes conferring disease resistance (Ozias-Akins and Gill, 2001; Magbanua et al., 2000; Yang et al., 1998; Higgins et al., 2004; Dar et al., 2006). Efficiency levels remain low and the process takes many months from when the initial transformation event is induced until plant maturity (Egnin et al., 1998). A high-efficiency, rapid technique to transform peanut is greatly needed and Agrobacterium-mediated transformation offers the possibility to achieve this goal. Cheng et al. (1996) used this method on a Valencia-type peanut, but other investigators have been unable to expand the methodology to other genotypes, thus restricting its usefulness. To date, biolistic methodologies are more reliable in peanut than other transformation methodologies and single constructs can be inserted into the peanut genome. Additional research is needed to insert multiple constructs and BAC clones to produce stable progenies. The regulatory process of germplasm release for consumption also must be explored as well as industry acceptance of transformed products.
Gene Sequencing in Arachis
Because the peanut is a polyploid with a large genome size, complete sequencing will be too expensive and labor intensive to perform with current resources. However, expressed sequence tags (ESTs) can give significant information about the function of genes. As of June, 2007, 12,832 long-sequence ESTs have been deposited in GenBank, with the majority from seeds. A large number of ESTs have been produced from seeds and leaves which will deposited in GenBank (Guo et al., 2004, 2008; Stalker and N. Nielson, unpublished data; Chen et al., 2006), and more than 25,000 unigenes have been identified from existing datasets (S. Knapp, personal commun.). In addition, Jayshree et al. (2005) identified 1312 short sequences that were isolated from SSR-enriched libraries and (S. Knapp, pers. commun.) is sequencing large numbers of short DNA segments with methylation-filtration techniques.
In addition to developing long and short read ESTs, several investigators have sequenced specific genes found in peanut. Two examples relevant to peanut are the isolation and characterization of the Δ12-fatty acid desaturase gene by Lopez et al. (2000) and sequencing the Ara h 2 gene which gives rise to one of the proteins causing allergens in humans by Ramos et al. (2006). These genes were sequenced to produce very specific markers to detect the genes in breeding or TILLING populations.
Genomic sequencing and microarray-based screening has been used to identify putative genes that may be associated with resistance to Aspergillus parasiticus and drought stress (Luo et al., 2005b) and for aflatoxin contamination (Guo et al., 2003). In addition, Yüksel et al. (2005) evaluated bacterial artificial clones from the library and found 250 putative resistance gene loci in peanut.
Strategic Planning – Peanut and Other Legumes
High-throughput genomic analyses are expensive and the vast majority of the financial resources for plants have been directed toward model systems. Recognizing the problem that significantly more resources will be required to make progress in agronomic plants (specifically in legumes) a landmark meeting was organized as the U.S. Legume Crops Genomics Workshop during 2001. Scientists involved in genomic research on soybean, peanut, common- and dry beans, alfalfa, peas and lentils, and model legumes met to develop a set of goals and strategy to present the case for increasing support for legume genomic research. They took a very broad view of genomics to include transformation-related research and recognized that molecular biology needs to be integrated with plant breeding efforts to be of long-term value. The outcome of the meeting was the development of a common set of goals across legume species in six areas of research, including: (a) genome sequencing of strategic legume species, (b) physical map development and refinement, (c) functional analysis including both transcriptional and genetic, (d) develop DNA markers for comparative mapping, (e) characterization and utilization of legume biodiversity, and (f) development of a legume data resource. Significantly, as a direct result of the meeting, the Legume Genomics Steering Committee was formed to organize the U.S. Legume Crops Genomics Initiative (USLCGI) to develop a strategy for additional funding for legumes. Members of this committee consisted of leaders in the United Soybean Board, American Soybean Council; National Peanut Foundation, American Peanut Council; National Dry Bean Council; USA Dry Pea and Lentil Council; Alfalfa and Clovers; Model Legumes; a USDA-ARS representative, and a researcher associated with each crop. National Research Initiative funding directed to legumes in 2005 was believed to be a result of their efforts. Additional outcomes of this committee was the book Legume Crop Genomics (Wilson et al., 2004) which documented research strategies, development of genomic tools, resources in legumes, and the legume-community consensus on the genomic research objectives.
The conference Legumes as a Model Plant Family: Genomics for Food and Feed, was held during 2004 in Santa Fe, NM. At this conference, 50 researchers developed a plan for genomic research with model species and across the legume crops alfalfa (Medicago sativa L.), common bean (Phaseolus vulgaris L.), pea (Pisum sativum L.), lentil (Lens culinaris Med.), chickpea (Cicer arietinu L. Gram.), peanut, and soybean (Glycine max L. Merr.). At the time of the conference, M. truncatula and Lotus japonicus (Regel) K. Larsen had been studied extensively as model species in the legume family (and soybean to a lesser extent), but other legume species were lagging behind in data generation and analyses (VandenBosch and Stacey, 2003). The goal of the conference was to develop a plan with specific objectives for cross-legume genomics research and to outline milestones for achieving these objectives (http://catg.ucdavis.edu; Gepts et al., 2005). A list of websites related to legumes is presented in Table 2. In addition to being economically important, peanut was of scientific interest to scientists working with other species because it is unique among cultivated legumes in mode of reproduction, is phylogenetically distinct, and has a unique nodule type.

Species-specific information resources.
To follow-up on the cross-legume workshop, on March 23, 2004, The Peanut Foundation/American Peanut Council hosted 26 scientists to document the current status of peanut genomic research and then establish priorities for the next 5 yr. Their efforts led to a Strategic Plan for Peanut Genomics 2004 to 2008 and an accompanying action plan to complete research (http://www.peanutbioscience.com/) that outlines priorities and milestones for peanut genomic research. The scientists took a very broad viewpoint of genomic technologies to include (a) simple markers to high-through put analysis, (b) gene discovery, (c) translational genetics, (d) transformation technologies, and (e) a recognition of the importance of germplasm and populations. The group set strategic goals to: (a) improve the utility of genetic tools for peanut genomics research, (b) improve the efficacy of technology for gene manipulation in genomics, (c) develop a framework for assembling the peanut genetic blueprint, (d) improve knowledge of gene identification and regulation, and (e) provide bioinformatics management of peanut biological information (http://www.peanutbioscience.com/). They also stated that a priority area for research in peanut is to increase peanut safety, especially by eliminating peanut allergens.
Summary of Progress in Peanut Genomic Research
Molecular markers should be useful for mapping and genetic studies, for estimating associations with plant and agronomic traits important to plant breeding efforts, to insure genes have been transferred from Arachis species to A. hypogaea in interspecific hybrid populations, to determine evolutionary relationships among species, and for plant identity assurances. The molecular data analyzed to date indicates that (a) variation within A. hypogaea ranges between 1 and 5% for different marker systems, (b) a large amount of variation exists among Arachis species for all marker systems, (c) very low-density maps have been created, (d) few genes of agronomic importance have been associated with various molecular markers, (e) introgression from Arachis species to A. hypogaea is possible, (f) transformation technologies are available for inserting genes from distantly related species to peanut, and (g) new molecular technologies promise to solve many of “low genetic variability” problems in peanut. To have effective use of genomic technologies in peanut (and other crop species), cooperative efforts must be developed between the molecular biologist and plant breeder to answer relevant biological questions and to utilize the tools made available by genomics. Successful utilization of molecular genetics for crop improvement will require integration of genomic science with classical methodologies to investigate problems that are too complex to be solved by traditional methodologies.
Literature Cited
Bianchi-Hall C. , Keys R. D. , Stalker H. T. , and Murphy J. P. 1993 Diversity of seed storage proteins in wild peanuts (Arachis species). Pl. Syst. Evol 186 : 1 – 15 .
Burks W. , Sampson H. A. , and Bannon G. A. 1998 Peanut allergens. Allergy 53 : 725 – 730 .
Burow M. D. , Simpson C. E. , Paterson A. H. , and Starr J. L. 1996 Identification of peanut (Arachis hypogaea L.) RAPD markers diagnostic of root-knot nematode (Melooidogyne arenaria (Neal) Chitwood) resistance. Molec. Breed 2 : 369 – 379 .
Chen H. , Holbrook C. C. , and Guo B. Z. 2006 Peanut seed transcriptome: construction of six peanut seed cDNA libraries from two peanut cultivars. Amer. Peanut Res. Educ. Soc 38 : 73 (abstr.) .
Chenault K. D. and Maas A. L. 2006 Identification of a simple sequence repeat (SSR) marker in cultivated peanut (Arachis hypogaea L.) potentially associated with Sclerotinia blight resistance. Proc. Amer. Peanut Res. Educ. Soc 37 : 24 – 25 (abstr.) .
Cheng M. , Jarret R. , Li Z. , Xing A. , and Demski J. 1996 Production of fertile transgenic peanut (Arachis hypogaea L.) plants using Agrobacterium tumefaciens. Plant Cell Rep 15 : 653 – 657 .
Chu Y. , Holbrook C. C. , Timper P. , and Ozias-Akins P. 2007 Development of a PCR-based molecular marker to select for nematode resistance in peanut. Crop Sci 47 : 841 – 845 .
Dar W. D. , Reddy B. V. S. , Gowda C. L. L. , and Ramesh S. 2006 Genetic resources enhancement of ICRISAT-mandate crops. Cur. Sci 91 : 880 – 884 .
Dhillon S. S. , Rake A. V. , and Miksche J. P. 1980 Reassociation kinetics and cytophotometric characterization of peanut (Arachis hypogaea L.) DNA. Plant Physiol 65 : 1121 – 1127 .
Egnin M. , Mora A. , and Prakash C. S. 1998 Factors enhancing Agrobacterium tumefaciens-mediated gene transfer in peanut (Arachis hypogaea L.). In Vitro Cell Dev. Biol. Plant 34 : 310 – 318 .
Ferguson M. E. , Burow M. D. , Schulze S. R. , Bramel P. J. , Paterson A. H. , Kresovich S. , and Mitchell S. 2004 Microsatellite identification and characterization in peanut (A. hypogaea L.). Theor. Appl. Genet 108 : 1064 – 1070 .
Garcia G. M. , Stalker H. T. , and Kochert G. A. 1995 Introgression analysis of an interspecific hybrid population in peanuts (Arachis hypogaea L.) using RFLP and RAPD markers. Genome 38 : 166 – 176 .
Garcia G. M. , Stalker H. T. , Shroeder E. , and Kochert G. 1996 Identification of RAPD, SCAR and RFLP markers tightly linked to nematode resistance genes introgressed from Arachis cardenasii to A. hypogaea. Genome 39 : 836 – 845 .
Garcia G. M. , Stalker H. T. , Schroeder E. , Lyerly J. H. , and Kochert G. 2005 A RAPD-based linkage map of peanut based on a backcross population between the two diploid species Arachis stenosperma and A. cardenasii. Peanut Sci 32 : 1 – 8 .
Gepts P. , Beavis W. D. , Brummer E. C. , Shoemaker R. C. , Stalker H. T. , Weeden N. F. , and Young N. D. 2005 Legumes as a model plant family. Genomics for food and feed report of the cross-legume advances through genomics conference. Plant Phys 137 : 1228 – 1235 .
Guo B. Z. , Yu J. , Lee R. D. , Holbrook C. C. , and Lynch R. E. 2003 Application of differential display RT-PCR and EST/microarray technologies to the analysis of gene expression in response to drought stress and elimination of aflatoxin contamination in corn and peanut. J. Toxicology-Toxin Reviews 22 : 291 – 316 .
Guo B. Z. , Chen X. , Dang P. , Scully B. T. , Liang X. , Holbrook C. C. , Yu J. , and Culbreath A. K. 2008 Peanut gene expression profiling in developing seeds at different reproduction stages during Aspergillus parasiticus infection. BMC Developmental Biology 8 : 12 .
Guo B.
,
Luo M.
,
Dang P.
,
He G.
, and
Holbrook C. C.
2004
Peanut expressed sequence tag (EST) project and the marker development for cultivated peanut (Arachis hypogaea).
Proc. International Crop Sci. Congress
2004
.
.
Halward T. , Stalker T. , LaRue E. , and Kochert G. 1992 Use of single-primer DNA amplifications in genetic studies of peanut (Arachis hypogaea L.). Plant Mol. Biol 18 : 315 – 325 .
Halward T. , Stalker H. T. , and Kochert G. 1993 Development of an RFLP linkage map in diploid peanut species. Theor. Appl. Genet 87 : 379 – 384 .
Halward T. , Stalker H. T. , Laure E. A. , and Kochert G. 1991 Genetic variation detectable with molecular markers among unadapted germplasm resources of cultivated peanut and related wild species. Genome 34 : 1013 – 1020 .
He G. and Prakash C. S. 1997 Identification of polymorphic DNA markers in cultivated peanut (Arachis hypogaea L.). Euphytica 97 : 143 – 149 .
He G. H. , Meng R. , Gao H. , Guo B. , Gao G. , Newman M. , Pittman R. N. , and Prakash C. S. 2005 Simple sequence repeat markers for botanical varieties of cultivated peanut (Arachis hypogaea L.). Euphytica 142 : 131 – 136 .
He G. H. , Meng R. , Newman M. , Gao G. , Pittman R. N. , and Prakash C. S. 2003 Microsatellites as DNA markers in cultivated peanut (Arachis hypogaea L.). BMC Plant Biology 3 : 3 .
Herselman L. 2003 Genetic variation among Southern African cultivated peanut (Arachis hypogaea L.) genotypes as reveled by AFLP analysis. Euphytica 133 : 319 – 327 .
Herselman L. , Thwaites R. , Kimmins F. M. , Courtois B. , van der Merwe P. J. A. , and Seal S. E. 2004 Identification and mapping of AFLP markers linked to peanut (Arachis hypogaea L.) resistance to the aphid vector of groundnut rosette disease. Theor. Appl. Genet 109 : 1426 – 1433 .
Higgins C. , Hall R. , Mitter N. , Cruickshank A. , and Dietzgen R. 2004 Peanut stripe potyvirus resistance in peanut (Arachis hypogaea L.) plants carrying viral coat protein gene sequences. Transgenic Res 13 : 59 – 67 .
Hilu K. and Stalker H. T. 1995 Genetic relationships between peanut and wild species of Arachis section Arachis (Fabaceae): Evidence from RAPDs. Plant Syst. Evol 188 : 167 – 178 .
Holbrook C. C. and Stalker H. T. 2003 Peanut breeding and genetic resources. Plant Breed. Rev 22 : 297 – 356 .
Hopkins M. S. , Casa A. M. , Wang T. , Mitchell S. E. , Dean R. E. , Kochert G. D. , and Kresovich S. 1999 Discovery and characterization of polymorphic simple sequence repeats (SSRs) in peanut. Crop Sci 39 : 1243 – 1247 .
Jain A. K. , Basha S. M. , and Holbrook C. C. 2001 Identification of drought - responsive transcripts in peanut (Arachis hypogaea L.). Mol. Biotechnology Genetics 4 / 2 :
Jayashree B. , Ferguson M. , Ilut D. , Doyle J. , and Crouch J. H. 2005 Analysis of genomic sequences from peanut (Arachis hypogaea). Elec. J. Biotech 8 : 226 – 237 .
Kochert G. , Halward T. , Branch W. D. , and Simpson C. E. 1991 RFLP variability in peanut (Arachis hypogaea L.) cultivars and wild species. Theor. Appl. Genet 81 : 565 – 570 .
Kochert G. , Stalker H. T. , Gimenes M. , Galgaro S. L. , Lopes C. R. , and Moore K. 1996 RFLP and cytological evidence on the origin and evolution of allotetraploid domesticated peanut, Arachis hypogaea (Leguminosae). Amer. J. Bot 83 : 1282 – 1291 .
Koppelman S. J. , Wensing M. , Ertmann M. , Knulst A. C. , and Knol E. F. 2004 Relevance of Ara h1, Ara h2 and Ara h3 in peanut-allergic patients, as determined by immoglobulin E Western blotting, basophile-histamine release and intracultaneous testing: Ara h2 is the most important peanut allergen. Clin. Exper. Allergy 34 : 583 – 590 .
Lanham P. G. , Fennell S. , Moss J. P. , and Powell W. 1992 Detection of polymorphic loci in Arachis germplasm using random amplified polymorphic DNAs. Genome 35 : 885 – 889 .
Liang X. Q. , Luo M. , Holbrook C. C. , and Guo B. Z. 2006 Storage protein profiles in Spanish and runner market type peanuts and potential markers. BMC Plant Biol 6 : 24 .
Lin Y. R.
,
Draye X.
,
Qian X.
,
Ren S.
,
Zhu L.
,
Tomkins J.
,
Wing R. A.
,
Li Z.
, and
Paterson A. H.
2000
Fine-scale mapping and sequence-ready contig assembly in highly-duplicated genomes, using the BAC-RF method.
Nucl. Acids Res
2823
.
.
Lopez Y. and Burow M. D. 2004 Development and validation of CAPS markers for the high oleate trait in peanuts. Proc. Amer. Peanut Res. Educ. Soc 36 : 25 – 26 (abstr.) .
Lopez Y. , Nadaf H. L. , Smith O. D. , Connell J. P. , Reddy A. S. , and Fritz A. K. 2000 Isolation and characterization of the Δ12-fatty acid desaturase in peanut (Arachis hypogaea L.) and search for polymorphisms for the high oleate trait in Spanish market-type lines. Theor. Appl. Genet 101 : 1131 – 1138 .
Lu J. and Pickersgill B. 1993 Isozyme variation and species relationships in peanut and its wild relatives (Arachis L. — Leguminosae) Theor. Appl. Genet 85 : 550 – 560 .
Luo M. , Dang P. , Guo B. Z. , He G. , Holbrook C. C. , Bausher M. G. , and Lee R. D. 2005a Generation of expressed sequence tags (ESTs) for gene discovery and marker development in cultivated peanut. Crop Sci 45 : 346 – 353 .
Luo M. , Liang X. Q. , Dang D. , Holbrook C. C. , Bausher M. G. , Lee R. D. , and Guo B. Z. 2005b Microarray-based screening of differentially expressed genes in peanut in response to Aspergillus parasiticus infection and drought stress. Plant Sci 169 : 695 – 703 .
Magbanua Z. , Wilde H. , Roberts J. , Chowdhury K. , Abad J. , Moyer J. , Wetzstein H. , and Parrott W. 2000 Field resistance to tomato spotted wilt virus in transgenic peanut (Arachis hypogaea L.) expressing an antisense nucleocapsid gene sequence. Mol. Breed 6 : 227 – 236 .
Milla S. R. , Tallury S. P. , Stalker H. T. , and Isleib T. G. 2004 Identification of molecular markers associated with tomato spotted wilt virus in a genetic linkage map of Arachis kuhlmannii × A. diogoi. Proc Am Peanut Res and Educ Soc 36 : 27 (abstr.) .
Milla S. R. , Isleib T. G. , and Tallury S. P. 2005 Identification of AFLP markers linked to reduced aflatoxin accumulation in A. cardenasii-derived germplasm lines of peanut. Proc. Amer. Peanut Res. and Educ. Soc 37 : 90 (abstr.) .
Moretzsohn M. C. , Leoi L. , Proite K. , Guimarães P. M. , Leal-Bertioli S. C. M. , Gimenes M. A. , Martins W. S. , Valls J. F. M. , Grattapaglia D. , and Bertioli D. J. 2005 A micro satellite–based, gene–rich linkage map for the AA genome of Arachis (Fabaceae). Theor. Appl. Genet 111 : 1060 – 1071 .
Nelson A. , Samuel D. M. , Tucker J. , Jackson C. , and Stahlecker-Roberson A. 2006 Assessment of genetic diversity and sectional boundaries in tetraploid peanuts (Arachis). Peanut Sci 33 : 64 – 67 .
Ozias-Akins P. and Gill R. 2001 Progress in the development of tissue culture and transformation methods applicable to the production of transgenic peanut. Peanut Sci 28 : 123 – 131 .
Ozias-Akins P. , Schnall J. A. , Anderson W. F. , Singsit C. , Clemente T. E. , Adang M. J. , and Weissinger A. K. 1993 Regeneration of transgenic peanut plants from stably transformed embryogenic callus. Plant Sci 93 : 185 – 194 .
Paik-Ro O. G. , Smith R. L. , and Knauft D. A. 1992 Restriction fragment length polymorphism evaluation of six peanut species within the Arachis section. Theor. Appl. Genet 84 : 201 – 208 .
Paterson A. H.
,
Stalker H. T.
,
Gallo-Meagher M.
,
Burow M. D.
,
Dwivedi S. L.
,
Crouch J. H.
, and
Mace E. S.
2004
Genomics and genetic enhancement of peanut.
97
–
109
In
Wilson R. F.
,
Stalker H. T.
, and
Brummer C. E.
Genomics for Legume Crops
Amer. Oil Chemical Soc. Press
Champaign, IL
.
Raina S. N. and Mukai Y. 1999 Genomic in situ hybridization in Arachis (Fabaceae) identifies the diploid wild progenitors of cultivated (A. hypogaea) and related wild (A. monticola) peanut species. Plant Sys. Evol 214 : 1 – 4 .
Ramos M. L. , Fleming G. , Chu Y. , Akiyama Y. , Gallo M. , and Ozias-Akins P. 2006 Genomic and evolutionary context for conglutin genes in Arachis. Mol. Gen 275 : 578 – 592.
Singh K. P. , Raina S. N. , and Singh A. K. 1996 Variation in chromosomal DNA associated with the evolution of Arachis species. Genome 39 : 890 – 897 .
Singh A. K. , Krishnan S. S. , Mengesha M. H. , and Ramaiah C. D. 1991 Polygenetic relationships in section Arachis based on seed protein profiles. Theor. Appl. Genet 82 : 593 – 597 .
Smartt J.
and
Stalker H. T.
1982
Speciation and cytogenetics in Arachis.
21
–
49
In
Pattee H.
and
Young C.
Peanut Science and Technology
Amer. Peanut Res. Educ. Soc
Yoakum, TX
.
Stalker H. T. , Phillips T. G. , Murphy J. P. , and Jones T. M. 1994 Diversity of isozyme patterns in Arachis species. Theor. Appl. Genet 87 : 746 – 755 .
Stalker H. T. , Dhesi J. S. , and Kochert G. D. 1995 Variation within the species A. duranensis, a possible progenitor of the cultivated peanut. Genome 38 : 1201 – 1212 .
Stalker H. T. and Mozingo L. G. 2001 Molecular markers of Arachis and marker- assisted selection. Peanut Sci 28 : 117 – 123 .
Temsch E. M. and Greihuber J. 2000 Genome size variation in Arachis hypogaea and A. monticola re-evaluated. Genome 43 : 449 – 451 .
Temsch E. M. and Greihuber J. 2001 Genome size in Arachis duranensis: A critical study. Genome 44 : 826 – 830 .
Vanden Bosch K. A. and Stacy G. 2003 Summaries of legume genomics projects from around the globe. Community resources for crops and models. Plant Physiol 131 : 840 – 865 .
Wilson R. F. , Stalker H. T. , and Brummer C. E. 2004 Genomics for Legume Crops Amer. Oil Chemical Soc. Press Champaign, IL 362 .
Yang H. , Singsit C. , Wang A. , Gonsalves D. , and Ozias-Akins P. 1998 Transgenic peanut plants containing a nucleocapsid protein gene of tomato spotted wilt virus show divergent levels of gene expression. Plant Cell Rep 17 : 693 – 699 .
Yüksel B. and Paterson A. H. 2005 Construction and characterization of peanut HindIII BAC library. Theor. Appl. Genet 111 : 630 – 639 .
Yüksel B. , Estill J. C. , Schulze S. R. , and Paterson A. H. 2005 Organization and evolution of resistance gene analogs in peanut. Mol. Gen. Genomics 274 : 248 – 263 .
Notes
Author Affiliations
1 Professor, Professor, and Research Assistant Professor, respectively, Department of Crop Science, Box 7620, North Carolina State Univ., Raleigh, NC 27695.
2 Research Geneticist, U. S. Department of Agriculture, Agricultural Research Service, PO Box 748, Tifton, GA 31793.
*Corresponding author: Tom_Stalker@ncsu.edu