

Introduction
Cylindrocladium black rot (CBR) of peanut (Arachis hypogaea L.), caused by the soilborne fungus Cylindrocladium parasiticum Crous, M. J. Wingf. & Alfenas (Crous, 2002), was first found in 1965 in Georgia (Bell and Sober, 1966) and threatens peanut production throughout the southeastern United States (Bell et al., 1973; Harris and Beute, 1982). The disease causes peg, pod, and root necrosis. The region immediately behind root tips is the primary infection court. Aboveground symptoms of CBR can appear in the field as early as July, and include chlorosis and wilting of the main stem followed by complete wilting and death of the plant. Taproots and hypocotyls become blackened and necrotic, with necrosis terminating at ground level. Lateral roots are either blackened or completely decayed. Pod development is greatly reduced, and existing pods may be severely rotted (Bell and Sober, 1966; Johnston and Beute, 1975). Yield losses of approximately 250 to 450 kg/ha for each 10% increase in CBR incidence have been reported (Pataky et al., 1983a).
Cultural practices, fumigants, fungicides, and resistant peanut genotypes have been evaluated for CBR management. Crop rotation with nonhosts and sanitation are useful to reduce populations of microsclertotia in soil. However, C. parasiticum is capable of long-term survival in the absence of hosts, and it is considered biologically impractical to eradicate microsclerotia from infested fields (Bell et al., 1973; Black et al., 1984; Rowe et al., 1974). Preplant fumigation with metam-sodium has been used successfully for CBR control (Phipps, 1990), but it is expensive and requires significant management input. Therefore, identification and utilization of resistance to CBR have been emphasized in peanut breeding programs (Beute et al., 1976; Coffelt and Garren, 1982; Isleib et al., 1997, 2003; Wynne and Beute, 1983; Wynne et al., 1991).
Results from several screening tests have shown that spanish-type peanuts are the least susceptible to C. parasiticum, valencia-type peanuts the most susceptible, while virginia-type peanuts are intermediate (Hammons et al., 1981; Harris and Beute, 1982; Phipps and Beute, 1997). However, large differences have been observed in the susceptibility of cultivars within each market type (Wynne et al., 1975). NC 3033, a virginia-type whose pedigree includes spanish-types, was released in 1976 as the first CBR-resistant germplasm (Beute et al., 1976). The NC breeding program has released several partially resistant virginia-type cultivars including ‘NC 8C’ (Wynne and Beute, 1983), ‘NC 10C’ (Wynne et al., 1991), ‘NC 12C’ (Isleib et al., 1997), and ‘Perry’ (Isleib et al., 2003). Georgia-02C, the first runner peanut cultivar with moderate resistance to CBR, was recently released (Branch, 2003).
A reliable and rapid technique for identifying resistance to CBR is very important for screening large numbers of lines in a breeding program. Evaluations of CBR reaction have been based on disease incidence, severity, or microsclerotium production in naturally infested fields, and in inoculated microplots and greenhouse experiments (Coffelt and Garren, 1982; Green et al., 1983a & b; Pataky et al., 1983b). However, inconsistent disease reactions frequently occur in these evaluations (Green et al., 1983a & b; Pataky et al., 1983b). Field evaluations of CBR resistance are not only time consuming, but also have resulted in large error components in the analysis of variance (Coffelt, 1980; Coffelt and Garren, 1982). This is due to the non-uniform spatial distribution of microsclerotia (Culbreath et al., 1990; Hau et al., 1982; Pataky et al., 1983b). Variability is also related to the inoculum density-dependent nature of CBR resistance (Black et al., 1984; Harris and Beute, 1982; Hau et al., 1982). Although inoculum density is more uniform in greenhouse screening, results do not always agree with those from field testing (Pataky et al., 1983b). Additionally, most of the evaluations of CBR resistance have been in virginia-, spanish-, and valencia-type peanuts (Coffelt, 1980; Coffelt and Garren, 1982; Garren and Coffelt, 1976; Pataky et al., 1983b). Few studies have been done with runner-type peanut, which is the major market type grown in Georgia. Therefore, the objective of this study was to compare the efficiency of different methods for evaluating CBR resistance in the field and greenhouse using runner-type peanut genotypes with varying reactions to CBR.
Materials and Methods
Genotypes Evaluated
The CBR-resistant cultivar Georgia-02C and the susceptible cultivar C-99R (Gorbet and Shokes, 2002) were used as reference entries to evaluate screening techniques. The reference cultivars were compared to the runner-type peanut genotypes Georgia-01R (Branch, 2002), Georganic (Holbrook and Culbreath, 2008), DP-1 (Gorbet, 2003), and the breeding line C34-24-85 in a naturally infested field, in fields following plant inoculations, and in artificially infested soil in the greenhouse. Georgia-01R is a recently released cultivar, which has shown some resistance to CBR (Branch, 2002). DP-1 (Gorbet and Shokes, 2002) and Georganic have previously shown multiple resistances to several diseases, such as early leaf spot, late leaf spot, and tomato spotted wilt virus (TSWV) (Cantonwine et al., 2006, 2008; Culbreath et al., 2005). C34-24-85 is a promising breeding line that was the most susceptible to CBR in preliminary tests.
Inoculum Production
Isolates CBR041, CBR0410, CBR0414, and CBR0418 of C. parasiticum, were obtained from naturally infected peanut plants in southern Georgia in 2004, were used to produce inoculum for field and greenhouse experiments. To obtain microsclerotia, the isolates were grown on potato-dextrose agar (PDA) for 6–7 wk, after which the cultures of each isolate were comminuted separately in a Waring Blender for 2 min and passed through nested sieves. Microsclerotia with in size the range of ≥53 to <425 µm were rinsed into small beaker and suspended in water.
Greenhouse Inoculation Technique
The two cultivars (Georgia-02C and C-99R) were grown in all possible combinations of four microsclerotial size ranges (≥53 to <75 µm, ≥75 to <150 µm, ≥150 to <250 µm, and ≥250 to < 425 µm) and five inoculum densities. The microsclerotia of each isolate were separated with five nested sieves with 425, 250, 150, 75, and 53 µm openings. Microsclerotia retained on the 425 µm sieve were discarded. The numbers of microsclerotia collected on each of the other four sieves were determined, and equal numbers of microsclerotia from each isolate were combined to give inoculum densities of 0, 0.5, 1.0, 5.0, and 10.0 microsclerotia/g soil. Appropriate amounts of suspensions were added to 3 kg premixed PRO-MIX ’BX’ (Premier Tech Ltd, Canada) and Robin Hood top soil(Robin Hood Garden & Landscape Products, GA) (1:1) in polyethylene bags. The infested soil was thoroughly mixed by shaking in bags for 2 min. Treatments were arranged in randomized complete block design. For each microsclerotial size × inoculum density combination, 7 replicate plastic cone-tainers (Stuewe & Sons, Inc., Corvallis, OR) (21 cm long by 3.8-cm diam.) with mesh on the bottom end were filled with infested or noninfested soil. A germinated seed was planted into each cone-tainer which was placed in a rack. The bottom one-third of the cone-tainers was submerged in water for the duration of the experiment to maintain a favorable environment for disease. Plants were harvested after growing for 8 wk in a greenhouse at ∼25°C. Root rot was visually estimated on a 0 to 5 scale, where 0 = no symptoms; 1 = some root discoloration, primarily on secondary roots; 2 = significant root browning and some necrosis, usually on secondary roots and tap root, with <25% of roots affected; 3 = moderate root rot, ≥25 to <75% of roots affected; 4 = severe root rot, ≥75% of roots affected; and 5 = dead plant. Crown rot ratings were assessed on a 0 to 3 scale, where 0 = no symptoms and 3 = completely rotted. This experiment was repeated once. Data for the root rot and crown rot rating scales were combined over the two trials. Analysis of variance was performed using the MIXED procedure of SAS (v.9.1, SAS Institute, Cary, NC). The effects of microsclerotial size and inoculum density were evaluated, and Fisher's least significant difference (LSD) values at α = 0.05 were computed using standard error and t values of adjusted degrees of freedom from the LSMEAN statement in Proc MIXED. The effect of inoculum density on root rot severity was evaluated with regression analysis for each cultivar using the REG procedures of SAS.
Evaluation of Genotypes in The Greenhouse
Based on size and density of microsclerotia that produced optimal level of disease, equal numbers of ≥150 to <250 µm microsclerotia from each of the four isolates were combined to infest soil at 1.0 microsclerotia/g soil as previously described. Seeds of each of the genotypes were planted in cone-tainers with infested soil, and noninfested soil as noninoculated controls. A randomized complete block design with six replicates per genotype, each containing seven plants was used. Root rot was assessed as described previously at 8 weeks after inoculation. Fresh plant weight and main stem height were also measured. Plant height reduction and whole plant weight reduction were calculated as a percentage of the non-inoculated control. This experiment was conducted twice. Data for root rot severity, disease incidence, plant height reduction, and plant weight reduction from the two trials were combined and analysis of variance was performed using the MIXED procedure of SAS. Genotype means were separated using Fisher's least significant difference (LSD) test at α = 0.05 computed using standard error and t values of adjusted degrees of freedom from the LSMEAN statement in Proc MIXED.
Evaluation of Genotypes by Inoculation of Plants in The Field
Plants were inoculated at two field sites in 2006 and 2007. Site 1 was at the Blackshank Farm in Tifton, GA, where the soil was a Tifton loamy sand (fine-loamy, siliceous, thermic plinthic kandiudults) previously planted to peanut. The field was tarped and fumigated with methyl bromide/chloropicrin (67/33) at 448 kg/ha 2 wk prior to planting. Genotypes were planted on 19 May 2006 and 16 May 2007. A split-plot design was used with four replications. Main plots were genotypes and subplots were inoculated or not with C. parasiticum. Subplots were two rows 0.91 m apart and 6.1 m long planted with 120 seeds per row. Entire subplots were inoculated at 50 days after planting with 480 ml of microsclerotia suspension containing 50 microsclerotia/ml with equal numbers of microsclerotia (150–250 and 250–425 µm) from each of the four isolates. A beaker was used to distribute the suspension of microsclerotia on the soil surface around the peanut plants. Overhead sprinklers then applied 2 cm of water to facilitate incorporation of the inoculum into the soil. Recommended cultural practices were followed for land preparation, fertilization and control of weeds and insects. Leaf spots caused by Cercospora arachidicola and Cercosporidium personatum were controlled by regular foliar applications of chlorothalonil 1.2 kg a.i. /ha (Bravo Ultrex®, Syngenta). Stem rot, caused by Sclerotium rolfsii, was controlled by an application of flutolanil at1.57 kg a.i./ha (Moncut® 70-DF, Gowan) at 60 days after planting. Numbers of diseased plants were counted before digging based on aboveground symptoms and after digging based on underground symptoms in each subplot. The total plant numbers per subplot were also counted after digging. The incidence of CBR above and underground were calculated as the percentage of plants with symptoms. Root rot severity was visually estimated on a 0 to 5 scale as described previously on ten plants randomly sampled from each inoculated subplot. The taproots were collected, and three pieces from each were surface sterilized in 0.25% sodium hypochlorite and plated on PDA to detect C. parasiticum. Colonies of C. parasiticum were counted after 5 days incubation at 25°C. Peanut pods were harvested from each plot separately. Pods were weighed after drying to 10% moisture. Yield reduction was calculated from yields of inoculated subplots expressed as a percentage of non-inoculated subplots for each genotype. A sample (500 g) of pods was collected from every inoculated subplot and the percentages of pod with rot were calculated. The interactions of year with disease incidence based on aboveground symptoms and underground symptoms, root rot severity, and percentage of pods with rot were significant (P < 0.05). Therefore, data from each of the two years were subjected to analysis of variance using the GLM procedure of SAS. Means were separated using Fisher's Protected LSD (P ≤ 0.05) Pearson's correlation coefficients among disease incidence based on aboveground symptoms and underground symptoms, root rot severity, isolation frequency, percentage of pods with rot, and percentage of yield reduction were computed by CORR procedure of SAS.
Site 2 was on the Tifton campus of the University of Georgia, where 170 × 140 cm2 microplots were filled with Tifton loamy sand (fine-loamy, siliceous, thermic plinthic kandiudults) to a depth of 100 cm. Each microplot was fumigated with metam sodium (Vapam) at 9.35 ml/m2 two weeks prior to planting on 16 May 2006 and 23 May 2007. Microplots contained two 140-cm long rows spaced 85 cm apart and planted with 25 seeds per row. A split-plot design was used with genotype as the main plot treatment and inoculation as the subplot treatment. A subplot row of every main plot was inoculated by applying a suspension of microsclerotia at 100 microsclerotia/plant to 8 plants/subplot 50 days after planting, while the other row was kept uninoculated. A stick (approximately 0.8-cm-dia.) was used to make two 5-cm deep holes around each inoculated plant, and 5 ml of a microsclerotia suspension (10 microsclerotia/ml) were added to each hole. Flags were used to mark the inoculated plants. Chlorothalonil at 1.2 kg a.i./ha was applied to foliage at 2- to 3-wk intervals to control leaf spot. Plots were drench-irrigated as needed to maintain soil moisture at favorable levels. Plants were individually dug with a shovel. The total number of diseased plants in each subplot was counted after digging, and the inoculation success rate in every subplot was calculated by dividing the number of diseased plants by the number of inoculated plants. Root rot was rated on each inoculated plants using the 0 to 5 scale described previously. Pods were picked by hand and yield was determined after drying to about 10% moisture for every subplot. The percentage of yield reduction was obtained by comparing the yields of the inoculated and non-inoculated subplots of each genotype. Taproots of all inoculated plants in each subplot were collected and plated as described previously. Data were analyzed as described for site 1.
Evaluation of Genotypes in Naturally Infested Fields
A field at the Southwest Research and Education Center in Plains, GA was used for evaluating cultivar resistance to CBR in 2005 and 2006. The field was a Greenville sandy clay loam soil and was planted to cotton in 2004, but previously was planted to peanut and had a history of severe CBR epidemics. Entries were replicated four or five times in two-row by 4.6-m plots with 80 seeds per row in 2005 and 2006. Rows were 0.91 m apart, and cultivars were arranged in a randomized complete block design. Crop management practices were the same as that used in field trials with inoculated plants.
CBR incidence based on both aboveground and underground symptoms, and root rot severity were assessed as described previously for site 1. All peanuts were picked by a combine and weighed after digging to approximately 10% moisture. Data were analyzed as described previously.
Results
Greenhouse Inoculation Technique
In greenhouse experiments, C. parasiticum mainly caused root tips to slough off and taproots to be black and necrotic. The mean root rot severity over microsclerotial and inoculum density on the susceptible genotype C-99R (2.03) was higher (P ≤ 0.05) than that on the partially resistant genotype Georgia-02C (1.27), which validated previous work. Obvious crown rot symptoms were also observed at harvest, but there were no differences in ratings over microsclerotial and inoculum density between C-99R (1.32) and Georgia-02C (1.24). Root rot is a more typical field symptom than crown rot, and this rating was apparently a better indicator of CBR resistance in the greenhouse screening.
The size of microsclerotia, inoculum level, genotype, and inoculum level × genotype effects on root rot severity were significant (P ≤ 0.05). The other two-way interactions and the three-way interaction were not significant (P > 0.05). The size of microsclerotia had significant effects on root rot severities on both resistant and susceptible peanut genotypes (Fig. 1). Generally, root severity increased with microsclerotial size. The large microsclerotia (≥250 to <425 µm) caused high root rot severity on the resistant genotype Georgia-02C at high inoculum densities (5 to 10 microsclerotia/g soil) that did not differ from the susceptible cultivar C-99R (Table 1). The ≥250 to <425-µm microsclerotia caused higher (P ≤ 0.05) root rot severities than the ≥150 to <250-µm microsclerotia at 5 to 10 microsclerotia/g soil on both C-99R and Georgia-02C (Table 1). Root rot severity for the moderately resistant genotype Georgia-02C and the susceptible C-99R differed in only three of the sixteen combinations of size of microsclerotia × inoculum density (Table 1). The use of the smaller microsclerotia (≥53 to <75 µm and ≥75 to <150 µm) did not result in genotype differences at any of the inoculum densities (data not presented). The effects of inoculum density were analyzed by genotype using the means of root rot severity across the size range of ≥150 to <250 µm and ≥250 to <425 µm of microsclerotia for the two greenhouse experiments using regression analysis. The linear effect of inoculum density was significant on root rot severity for both resistant (r2 = 0.90, P = 0.01) and susceptible (r2 = 0.57, P = 0.04) genotypes. Generally, the root rot severity increased as the inoculum density increased. The regression line slopes for the two genotypes were similar (P > 0.05), but the intercept for C-99R was greater than that for Georgia-02C (P < 0.01) (Fig. 2)
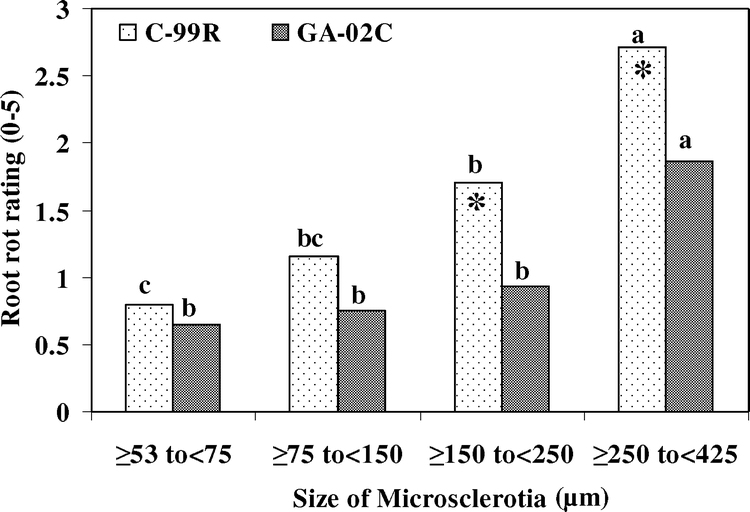
Effects of size of microsclerotia (across all densities of 0 to 10 microsclerotia/g soil) on root rot severity in two peanut genotypes C-99R and Georgia-02C in the greenhouse inoculations. Means within the same genotype followed by the same letter are not different (P > 0.05), according to Fisher's least significant difference t test. (*) = means differed between cultivars at P ≤ 0.05 within a microsclerotial size range. Root rot severity on a scale of 0 to 5, where 0 = no visible disease symptoms and 5 = completely decayed. CK = no inoculum.
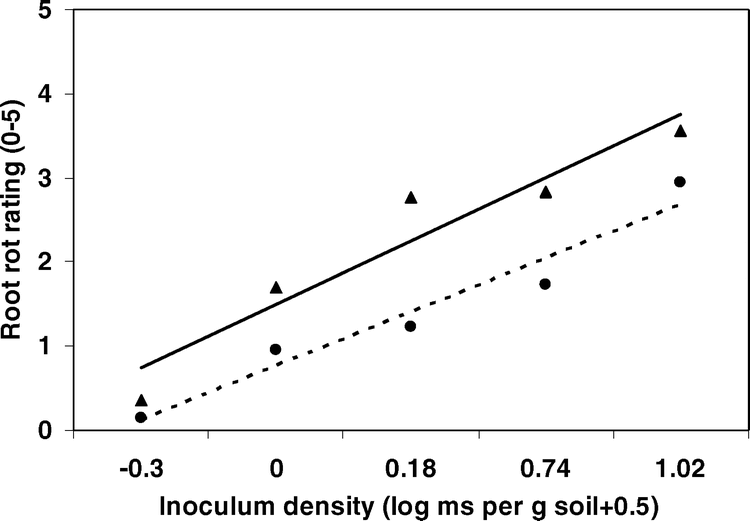
Effects of inoculum density of microsclerotia in the size range of >150 to <425 µm on root rot severities in two peanut genotypes C-99R and Georgia-02C in greenhouse inoculations. Data points are means of root rot severity of two trials (seven replications/trial). Root rot severity on a scale of 0 to 5, where 0 = no visible disease symptoms and 5 = completely decayed. (——▴——) = C-99R, y = 1.57 + 0.90x (r2 = 0.75, P = 0.04); (-----•-----) = GA-02C, y = 0.80 + 0.80x (r2 = 0.90, P = 0.01).
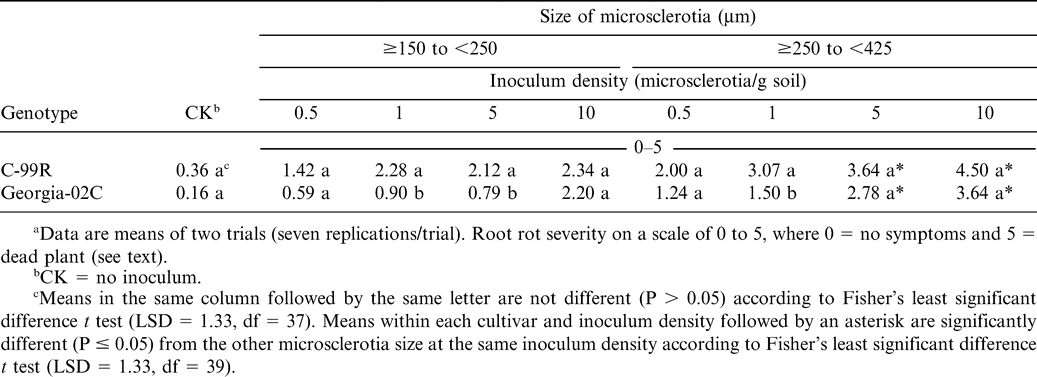
Root rot severitya for two peanut genotypes (Georgia-02C and C-99R), in response to size and density of Cylindrocladium parasiticum microsclerotia in greenhouse trials.
Evaluation of Genotypes in The Greenhouse
At 8 weeks after inoculation, CBR incidence for all genotypes was over 90%. However, disease incidence for Georgia-02C was lower (P < 0.05) than for the other genotypes (Table 2). Georgia-02C also had less root rot than the other genotypes, while DP-1 had lower root rot severity than the most susceptible genotypes, C34-24-85 and Georgia-01R.
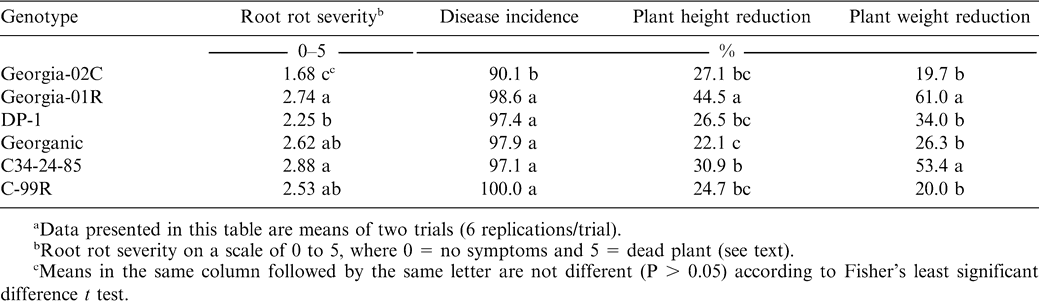
Responses of peanut genotypes to inoculation with Cylindrocladium parasiticum in the greenhouse.a
For all the genotypes, CBR infection reduced plant main stem height and plant fresh weight compared to non-inoculated plants (Table 2). The reference genotypes C-99R and Georgia 02C had similar reductions in plant height and plant weight. The greatest impact on main stem height was for Georgia-01R, which was 44.5% shorter than the average of plants without CBR. Georganic showed less reduction in plant height than C34-24-85. The genotypes Georgia-01R and C34-24-85 also showed the highest percentage weight reductions from CBR at 61.0% and 53.4%, respectively. There was a positive correlation between root rot severities and reductions in both height (r = 0.34, P = 0.04) and weight (r = 0.58, P < 0.0001). Root rot was also correlated with CBR incidence (r = 0.45, P < 0.0001); however, disease incidence was high (> 90%) for all genotypes and not correlated with plant height or weight.
Evaluation of Genotypes by Inoculation in The Field
At site 1, the noninoculated subplots were CBR free. Thus, only the disease severity data taken from inoculated subplots were used to assess the genotype reactions. However, both the inoculated and noninoculated subplot data were used to calculate the percent yield reduction from CBR. In both years, CBR incidence based on underground symptoms was significantly lower for Georgia-02C than for C-99R which showed the highest incidence among the six genotypes (Table 3). Except for Georganic and C34-24-85 in 2006, CBR incidences for the other genotypes were not significantly different from C-99R and Georgia-02C. The cultivars C-99R and DP-1 were among the genotypes with high values for CBR incidence based on aboveground symptoms and the highest root rot severities. Georgia-02C had fewer black pods than did Georganic or C-99R, but none of the other genotypes differed for pod disease. In both 2006 and 2007, the yield reduction and C. parasiticum isolation rates from the taproots of plants from the inoculated subplots did not differ among genotypes.
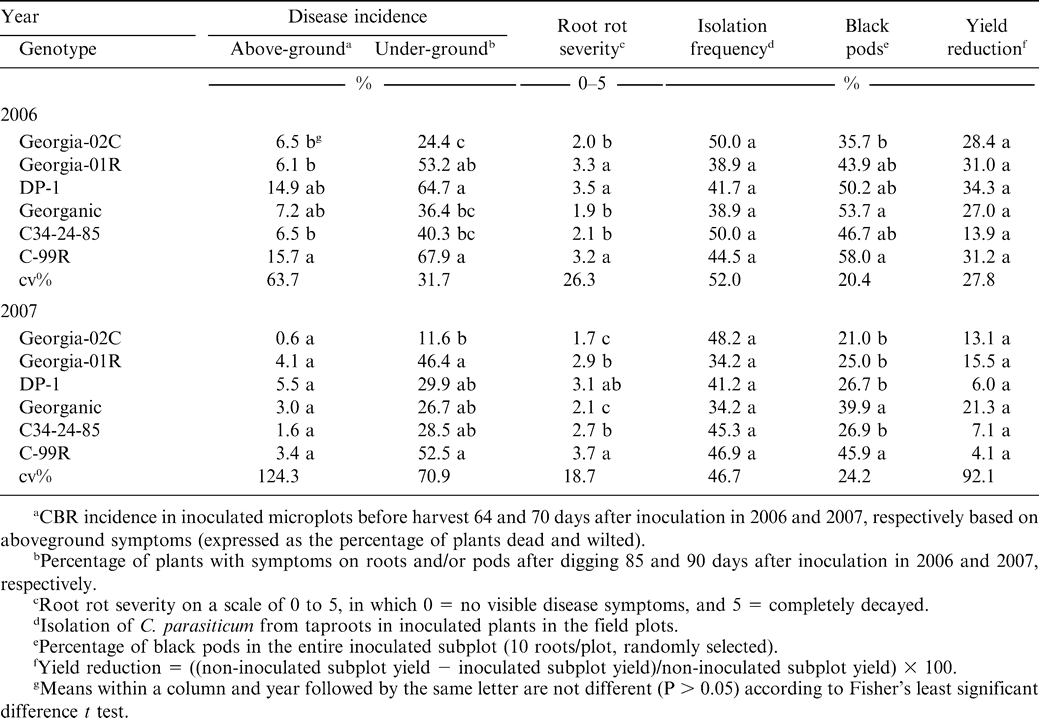
Cylindrocladium black rot (CBR) reactions, yield effects, and pod damage for different peanut genotypes inoculated with Cylindrocladium parasiticum in a fumigated field.
There were significant correlations (P < 0.01) between disease incidence based on both aboveground and underground symptoms, and root rot severity. The correlation coefficient between disease incidence based on underground symptoms and root rot severity (r = 0.69) was greater than that between disease incidence based on aboveground symptoms and root rot severity (r = 0.43). Disease incidence based on underground symptoms was correlated with the percentage of black pods (r = 0.45, P = 0.03), while disease incidence based on aboveground symptoms was not. Neither disease incidence based on aboveground symptoms nor disease incidence based on underground symptoms was correlated with yield reduction or C. parasiticum isolation frequency.
In the microplots with individually inoculated plants, one or more of the plants per inoculation site developed symptoms among the susceptible genotypes, but there was less than one diseased plant per inoculation site for the more resistant genotypes. The responses of genotypes to CBR inoculation were similar to those observed at site 1 (Table 3). The peanut cultivars Georgia-02C and Georganic showed higher resistance than cultivars C-99R and DP-1, with Georgia-01R being intermediate in both 2006 and 2007 (Table 4). The inoculation success rate of C34-24-85 was inconsistent between years. The root rot severities of the inoculated plants of genotypes Georgia-02C, Georganic, and C34-24-85 were numerically lower than those for C-99R and DP-1, but the differences were not always significant, particularly in 2006. Differences among genotypes for C. parasiticum isolation frequency at site 2 were not significant (Table 4).
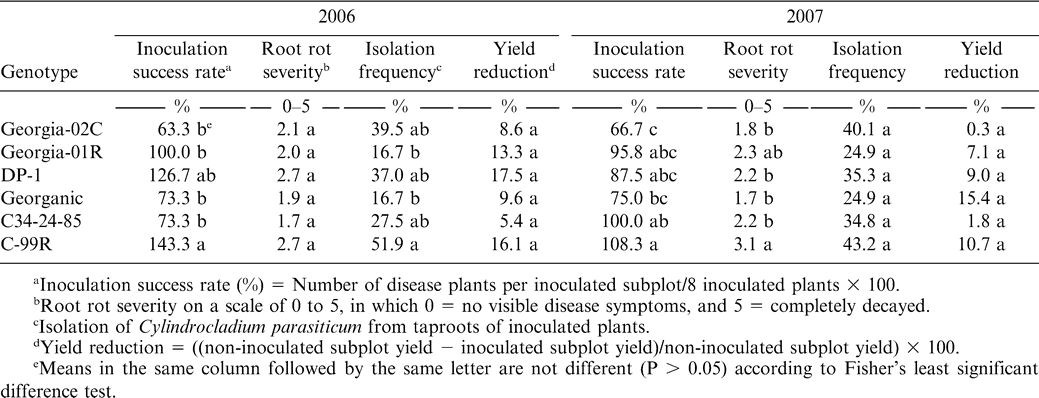
Cylindrocladium black rot (CBR) and yield response of peanut genotypes in microplots where individual plants were inoculated with Cylindrocladium parasiticum.
Root rot severity was significantly correlated with inoculation success rate and pathogen isolation frequency (r = 0.78, P ≤ 0.01). There were also significant correlations between isolation frequency, and both inoculation success rate (r = 0.54, P ≤ 0.05) and root rot severity (r = 0.62, P ≤ 0.01). However, neither inoculation success rate nor root rot severity was correlated with yield reduction (P > 0.05).
Evaluation of Genotypes in Naturally Infested Fields
Both root rot severity and incidence of CBR were greater in 2006 compared to 2005; however, pod yields of genotypes were not lower in 2006 than in 2005 (Table 5). Only C34-24-85 had a higher incidence of aboveground symptoms before harvest than other genotypes in 2005, whereas disease incidence before harvest for C-99R was higher than in the other genotypes in 2006. Incidence of CBR after harvest based on underground symptoms was higher in 2006 than 2005, but genotypes were similarly ranked in both years, except for C34-24-85. This genotype showed the greatest number of diseased plants and the highest root rot severity among the six genotypes in 2005; however, it was in the most resistant group of cultivars in 2006. In both years, the disease incidence after harvest and root rot severity for Georgia-02C were significantly lower than those of C-99R. In 2006, Georgia-02C also showed better resistance than Georgia-01R and DP-1 based on the root rot severity, while C-99R showed higher susceptibility than Georganic, GA-01R, and C34-24-85.

Cylindrocladium black rot (CBR) and yield responses of peanut genotypes in field plots naturally infested with Cylindrocladium parasiticum at Plains, GA in 2005 and 2006.
Disease incidence both before and after and the root rot severities in 2005 and 2006 were positively correlated with each other. The correlation coefficients of disease incidence after harvest and root rot severities were 0.80 (P < 0.01) and 0.78 (P < 0.01) in 2005 and 2006, respectively, and were higher than those for disease incidence before harvest and root rot severities in 2005 (r = 0.68, P < 0.01) and 2006 (r = 0.49, P < 0.05). Disease incidence after harvest also had a lower coefficient of variation (cv%) than disease incidence before harvest (Table 5), and was easier to assess than root rot severity.
Correlation of Field and Greenhouse Results
The ratings of disease incidence after harvest (underground symptoms) for the naturally infested field experiment were significantly correlated with those in the field experiments using inoculated plants (r = 0.84, P < 0.01), but neither was correlated with root rot severity for the greenhouse experiments (P > 0.05). There were no significant correlations observed between the percentage of yield reduction in naturally infested field experiments and root rot severity, disease incidence after harvest for inoculated plants in the field, or disease incidence after harvest in naturally infested fields (P > 0.05).
Discussion
Resistance to Cylindrocladium parasiticum in peanut is dependent upon inoculum density (Black and Beute, 1984; Phipps and Beute, 1977). High inoculum densities can cause serious damage even on partially resistant peanut genotypes. Our results showed that not only inoculum density, but also the size of microsclerotia were related to root rot severities. At the same inoculum density, root rot was more severe for large microsclerotia. This was consistent with previous reports (Black and Beute, 1984). The most effective inoculum density for detecting differences in the degree of susceptibility was 1–5 microsclerotia/g soil for microsclerotia in the 150–425 µm size range. Results of this study confirm that it is critical to standardize propagule size and density to obtain reproducible results for germplasm screening in the greenhouse.
In greenhouse experiments, some peanut plants showed obvious crown rot symptoms. Such symptoms usually occur in the field only after significant root disease. The crown rot phase of CBR represents a potentially different component of resistance so it was also evaluated. Some crowns were completely rotted, while the root systems were still relatively unaffected. However, crown rot ratings were not a good criterion for resistance identification, due to the lack of differences between genotypes. This phase of the disease is also seldom observed in the field apart from significant root damage and may be an artifact of greenhouse inoculations.
Resistance of plants to pathogens has been defined as the ability of the plant to lessen, inhibit, or overcome the attack by the pathogen (Wingard, 1953). However, in screening peanut for resistance to CBR, the host response to the pathogen is more meaningful. Researchers have used incidence of dead and diseased plants, root rot severity (Coffelt, 1980; Green et al., 1983b; Pataky et al., 1983b), and pod damage (Coffelt and Garren, 1982; Wynne et al., 1975) to screen for resistance to CBR in peanut. In this study, several components of disease development were assessed following inoculation of plants in the field, including CBR incidence based on aboveground and underground symptoms, root rot, and pod damage. Based on disease incidence, inoculation success rate, and root rot severity, the two field tests in which plants were inoculated had similar genotype responses. It is interesting to note that with the most susceptible genotype there was sometimes greater than 100% inoculation success (Table 4), indicating that some adjacent plants also developed symptoms. Presumably, this would have been from roots of those plants growing into the infested soil around the inoculated plant. The disease incidence based on underground symptoms, inoculation success rate and root rot severity were lower for Georgia-02C than for the susceptible cultivar C-99R, but the isolation frequencies of C. parasiticum from inoculated plants of Georgia-02C were similar to those from C-99R. This indicates that Georgia-02C may be infected as readily as C-99R by C. parasiticum, but apparently disease development was slower on Georgia-02C. We also evaluated the degree of yield reduction from CBR by comparing the yield of inoculated and noninoculated subplots of the same genotype, thus identifying those genotypes with tolerance to CBR (i.e. those which may produce good yield in spite of having high disease incidence and severity). However, there were no differences in yield reduction among the cultivars, suggesting a similar level of tolerance among genotypes even for the more susceptible lines. One apparent conclusion is that the resistance to CBR in the runner-type genotypes evaluated here is at best a partial resistance, and that more highly resistant cultivars are needed to effectively manage this disease.
We assessed the incidence of diseased and dead plants both before and after digging, and rated root rot on plants selected at random in the naturally infested field tests. Tomato spotted wilt virus (TSWV) can cause symptoms similar to those caused by CBR on peanut roots (Culbreath et al., 1991), and may confound root rot severity assessment. Therefore, we also evaluated incidence of TSWV based on the aboveground symptoms and analyzed the correlation coefficients between TSWV incidence and CBR incidence after digging and root rot severity. There were no significant correlations (data not shown), indicating that TSWV had little effect on disease incidence after digging and root rot severity evaluations in our field experiments. However, more severe epidemics of tomato spotted wilt than those experienced during this study may complicate both aboveground and below ground evaluation of CBR incidence and severity. Generally, disease incidence after digging is the best variable for screening a large number of peanut lines in the field for CBR resistance. If time permits, the root rot severity rating also should be assessed because more than one resistance mechanism may exist in peanut (Coffelt and Garren, 1982).
In the naturally infested and inoculated field experiments, and the greenhouse experiments, Georgia-02C and Georganic were among the genotypes with the highest resistance to CBR. Cultivars C-99R and DP-1 were the most susceptible in naturally infested and inoculated plants in the field experiments, but their root rot severities were not the highest in the greenhouse experiments. In summary, the results of naturally infested field tests and genotype inoculations in the field were consistent, but the greenhouse results were consistent only for the most resistant and most susceptible genotypes. Peanut genotypes were reliably screened by plant inoculations in the field or evaluation of genotypes in uniformly naturally infested fields. Greenhouse evaluations provide the faster results, but results were not well correlated with those from the field. Recent work on virginia-type peanuts showed that germplasm reactions from greenhouse inoculations were significantly correlated with field results (Hollowell et al., 2008). Therefore specific inoculation techniques may affect results and should be verified with runner-type peanuts to facilitate germplasm evaluation.
Literature Cited
Bell D. K. and Sobers E. K. 1966 A peg, pod, and root necrosis of peanut caused by a species of Calonectria. Phytopathology 56 : 1361 – 1364 .
Bell D. K. , Locke B. J. , and Thompson S. S. 1973 The status of Cylindrocladium black rot of peanut in Georgia since its discovery in 1965. Plant Dis. Rep 57 : 90 – 94 .
Beute M. K. , Wynne J. C. , and Emery D. A. 1976 Registration of NC 3033 peanut germplasm. Crop Sci 16 : 887 .
Black M. C. and Beute M. K. 1984 Relationships among inoculum density, microsclerotium size, and inoculum efficiency of Cylindrocladium crotalariae causing root rot on peanuts. Phytopathology 74 : 1128 – 1132 .
Black M. C. , Beute M. K. , and Leonard K. J. 1984 Effects of monoculture with susceptible and resistant peanuts on virulence of Cylindrocladium crotalariae. Phytopathology 74 : 945 – 950 .
Branch W. D. 2002 Registration of ‘Georgia-01R’ peanut. Crop Sci 43 : 1750 – 1751 .
Branch W. D. 2003 Registration of ‘Georgia-02C’ peanut. Crop Sci 43 : 1883 – 1884 .
Cantonwine E. G. , Culbreath A. K. , Holbrook C. C. , and Gobert D. W. 2008 Disease progress of early leaf spot and components of resistance to Cercospora arachidicola and Cercosporidium personatum in runner-type peanut cultivars. Peanut Sci 34 . (in press) .
Cantonwine E. G. , Culbreath A. K. , Stevenson K. L. , Kemerait R. C. , Brenneman T. B. , Smith N. B. , and Mullinex B. G. 2006 Integrated disease management of leaf spot and spotted wilt of peanut. Plant Dis 90 : 493 – 500 .
Coffelt T. A. 1980 Reaction of spanish-type peanut genotypes to Cylindrocladium black rot. Peanut Sci 7 : 91 – 94 .
Coffelt T. A. and Garren K. H. 1982 Screening for resistance to Cylindrocladium black rot in peanuts (Arachis hypogaea L.). Peanut Sci 9 : 1 – 5 .
Crous P. W. 2002 Taxonomy and pathology of Cylindrocladium (Calonectria) and allied genera The American Phytopathology Society St. Paul, Minnesota 91 – 94 .
Culbreath A. K. , Beute M. K. , and Wynne J. C. 1990 Use of spatial patterns and density of inoculum of Cylindrocladium crotalariae during field evaluation of partially resistant peanut genotypes. Phytopathology 80 : 1395 – 1400 .
Culbreath A. K. , Csinos A. S. , Brenneman T. B. , Todd J. W. , and Demski J. W. 1991 Association of tomato spotted wilt virus with general chorosis of peanut. Plant Dis 75 : 863 .
Culbreath A. K. , Gorbet D. W. , Martinez-Ochoa N. , Holbrook C. C. , Todd J. W. , Isleib T. G. , and Tillman B. L. 2005 High levels of field resistance to tomato spotted wilt virus in peanut breeding lines derived from hypogaea and hirsute botanical varieties. Peanut Sci 32 : 20 – 24 .
Garren K. H. and Coffelt T. A. 1976 Reaction to Cylindrocladium black rot in virginia-type peanut cultivars. Plant Dis. Rept 60 : 175 – 178 .
Gorbet D. W. and Shokes F. M. 2002 Registration of ‘C-99R’ Peanut. Crop Sci 42 : 2207 .
Gorbet D. W. 2003 New University of Florida peanut varieties for 2003. 03-2, Marianna NFREC Res. Rep., Marianna, FL.
Green C. C. , Beute M. K. , and Wynne J. C. 1983a A comparison of methods of evaluating resistance to Cylindrocladium crotalariae in peanut field tests. Peanut Sci 10 : 66 – 69 .
Green C. C. , Wynne J. C. , and Beute M. K. 1983b Genetic variability and heritability estimates based on the F2 generation from crosses of large-seeded virginia-type peanuts with lines resistant to Cylindrocladium black rot. Peanut Sci 10 : 47 – 51 .
Hammons R. O. , Bell D. K. , and Sobers E. K. 1981 Evaluating peanuts for resistance to Cylindrocladium black rot. Peanut Sci 8 : 117 – 120 .
Harris N. E. and Beute M. K. 1982 Histological responses of peanut germplasm resistant and susceptible to Cylindrocladium crotalariae in relationship to inoculum density. Phytopathology 72 : 1250 – 1256 .
Hau F. C. , Campbell C. L. , and Beute M. K. 1982 Inoculum distribution and sampling methods for Cylindrocladium crotalariae in a peanut field. Plant Dis 66 : 568 – 571 .
Holbrook C. C. and Culbreath A. K. 2008 Registration of ‘Georganic’ peanut. J. Plant Registr 2 : 17 .
Hollowell J. E. , Isleib T. G. , Tallury S. P. , Copeland S. C. , and Shew B. B. 2008 Screening of virginia-type peanut breeding lines for resistance to Cylindrocladium black rot and Sclerotinia blight in the greenhouse. Peanut Sci 35 : 18 – 24 .
Isleib T. G. , Rice P. W. , Bailey J. E. , Mozingo R. W. , and Pattee H. E. 1997 Registration of ‘NC 12C’ peanut. Crop Sci 37 : 1976 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Bailey J. E. , Mozingo R. W. , and Pattee H. E. 2003 Registration of ‘Perry’ peanut. Crop Sci 43 : 739 – 740 .
Johnston S. A. and Beute M. K. 1975 Histopathology of Cylindrocladium black rot of peanut. Phytopathology 65 : 649 – 653 .
Pataky J. K. , Beute M. K. , Wynne J. C. , and Carlson G. A. 1983a Peanut, yield, market quality and value reductions due to Cylindrocladium black rot. Peanut Sci 10 : 62 – 66 .
Pataky J. K. , Black M. C. , Beute M. K. , and Wynne J. C. 1983b Comparative analysis of Cylindrocladium black rot resistance in peanut: greenhouse, microplot, and field testing procedures. Phytopathology 73 : 1615 – 1620 .
Phipps P. M. 1990 Control of Cylindrocladium black rot of peanut with soil fumigants having methyl isothiocyanate as the active ingredient. Plant Dis 74 : 738 – 741 .
Phipps P. M. and Beute M. K. 1977 Sensitivity of susceptible and resistant peanut cultivars to inoculum densities of Cylindrocladium crotalariae microsclerotia in soil. Plant Dis. Reptr 61 : 300 – 303 .
Phipps P. M. and Beute M. K. 1997 Cylindrocladium black rot. 12 – 15 In: Kokalis-Burelle N. , Porter D. M. , Rodriguez-Kabana R. , Smith D. H. , and Subrahmanyam P. Compendium of peanut diseases. 2nd ed APS Press, Am. Phytopath. Soc St. Paul, MN .
Rowe R. C. , Beute M. K. , and Wynne J. C. 1974 Incidence and control of Cylindrocladium black rot of peanut in North Carolina during 1973. Plant Dis. Reptr 58 : 348 – 352 .
Wingard S. A. 1953 The nature of resistance to disease. 165 – 173 In: the Year-book of Agriculture US Department of Agriculture Washington, DC .
Wynne J. C. and Beute M. K. 1983 Registration of ‘NC 8C’ peanut. Crop Sci 23 : 183 – 184 .
Wynne J. C. , Beute M. K. , Bailey J. , and Mozingo R. W. 1991 Registration of ‘NC 10C’ peanut. Crop Sci 31 : 484 .
Wynne J. C. , Rowe R. C. , and Beute M. K. 1975 Resistance of peanut genotypes to Cylindrocladium crotalariae. Peanut Sci 2 : 54 – 56 .
Notes
- First and second authors: Res. Agron. , USDA-ARS and Agric. Res. Statistician; Coastal Plain Exp. Sta., Tifton, GA 31793-0748. Third author: President and CEO, Hebert Green AgroEcology; Asheville, NC 28801. [^]
- Department of Plant Pathology, University of Georgia, Coastal Plain Experiment Station, Tifton, GA 31793 [^]
- USDA-ARS, Coastal Plain Experiment Station, Tifton, GA 31793 [^] *Corresponding author: W. B. Dong (E-mail: wbdong@uga.edu)
Acknowledgements
This research was funded in part by the Georgia Peanut Commission. We thank Patricia Hilton for her help in many aspects of this study. We thank Jimmy Mixon, Lewis Mullis, Russell Griffin, Dannie Mauldin, Jason Golden, and Amber Graham for their field and lab assistance.
Author Affiliations