

Introduction
The use of biotechnology to produce transgenic crops is rapidly becoming a viable extension of conventional plant breeding, particularly since transgenic plants are now being produced to meet broad objectives such as pathogen and herbicide resistance. Genetic engineering promises to provide those involved in agricultural biology with the tools needed to overcome the limited gene pools accessible to conventional breeding programs. The list of crop species being transformed with genes responsible for agronomically beneficial traits is rapidly expanding. Herbicide resistance has been engineered in a wide variety of crops including, but not limited to, soybean (Delannay et al., 1995), tobacco (Brandle and Miki, 1993), cotton (Coulombe et al., 1994), sorghum (Casas et al., 1993), wheat (Vasil et al., 1992), sugar beet (D'Halluin et al., 1992), and alfalfa (D'Halluin et al., 1990). Engineered crop resistance to pathogen attack has also become common place. Virus resistance in squash (Arce-Ochoa et al., 1995; Tricoli, et al., 1995) and tomato (Fuchs and Provvidenti, 1996), bacterial resistance in potato (Allefs et al., 1995), fungal resistance in rice (Lin et al.,1995), and insect resistance in cotton (Wilson et al., 1994) are only a few examples of these efforts.
Peanut is susceptible to many pathogens, with most damage being caused by fungi (Melouk and Backman, 1995). Soilborne fungi cause diseases that adversely affect peanut health and productivity throughout the peanut growing areas of the United States. Diseases such as pod rot (Rhizoctonia solani Kühn, Pythium myriotylum), crown rot (Aspergillus niger Teigh), southern blight (Sclerotium rolfsii Sacc) and root knot (Meloidogyne arenaria [Neal] Chitwood) occur in all U.S. peanut-producing areas, while others such as Sclerotinia blight (Sclerotinia minor Jagger) are limited to certain geographic regions. Sclerotina blight is of major concern to peanut producers in the Southwest U.S. Traditional breeding and screening practices have resulted in few cultivars resistant to fungal diseases that are suitable for commercial use (Smith et al., 1991; Simpson et al., 2000), making expensive fungicide applications throughout the growing season required for effective disease management. These obstacles to profitable peanut production along with recent reductions in the U.S. peanut price support system have resulted in the urgent need for effective alternative methods of disease management that will provide disease resistance without pesticide application.
Efforts to engineer fungal resistance in peanut have resulted in the production of transgenic peanut lines that express a chitinase from rice (Oryza sativa L.) and or a glucanase from alfalfa (Medicago sativa L.) (Chenault et al., 2002). Pathogenesis-related (PR) proteins such as chitinases and β-1-3 glucanases that are capable of hydrolyzing the cell walls of many fungi that attack plants are rational candidates for over-expression to produce disease-resistant crops (Stahl and Bishop, 2000). The cultivar Okrun served as the parent genotype to all transgenic lines produced due to its desirable and marketable agronomic characteristics and its performance under tissue culture conditions. Resulting transgenic peanut lines were analyzed for stable transgene inheritance and expression (Chenault et al., 2002), as well as for their response to fungal infection under greenhouse conditions (Chenault et al., 2003).
The evaluation of transgenic plants in the laboratory and greenhouse provides valuable and necessary information about the inheritance and expression of the introduced gene(s). However, the introduction of transgene(s) into the plant genome is not always site-specific and endogenous genes that perform essential functions in plant productivity may be disrupted. True assessment of plant phenotype and performance can be determined only in a field environment with inherent environmental variations. Moreover, field testing of transgenic crops is an essential step toward variety release and commercial production. The purpose of this study was to determine whether transgenic peanut lines expressing anti-fungal genes have retained the desirable agronomic characteristics of the parent genotype Okrun under field conditions.
Materials and Methods
Transgenic peanut lines were produced from somatic embryos of the cultivar Okrun and analyzed as previously reported (Yang et al., 1998; Chenault et al, 2002). Thirty-two transgenic lines with single-copy transgene insertions were chosen for evaluation in field experiments. Field tests were conducted for three growing seasons (2000, 2001 and 2002) at the Oklahoma State University Agricultural Experiment Station in Caddo County, OK. Disease inoculum was assessed for each test by determining the number of S. minor sclerotia present in soil samples taken from each plot. Viable sclerotial density was determined in the top 5 cm of field soil by a modified elutriation technique (Porter and Steele, 1983). Each individual plot was 6.1 m × 7.6 m and consisted of six rows (0.91 m row spacing): four rows of transgenic test lines, one of the resistant cultivar Southwest Runner (Kirby et al., 1998), and one of the susceptible cultivar Okrun. Plots were arranged in a randomized complete block design with 8 plots/replication and 3 replications per test. The presence of transgene(s) is transgenic lines was confirmed via PCR as previously reported (Chenault et al., 2002) before planting. All kernels were treated with TOPS 90 fungicide (2.5 g/kg kernel) before planting. Transgenic kernels to be planted the first year of evaluation (2000) originated under greenhouse conditions (T5), but were taken from the field plots for the two subsequent years (T6 and T7). Kernels were hand-planted 23 cm apart on the 15th of May, 2000–2002.
All plots were weeded on a weekly basis and assessed for disease symptoms. Disease incidence (#diseased plants/line/replication) was recorded at weekly intervals after initial onset. The percentage of plants infected by Sclerotinia blight was determined by the presence of visible above-ground symptoms. Individual plants were considered infected if symptoms were present, however slight or severe, and marked with a flag at the time of recording. Disease data were tested for statistical significance with SAS v. 8.2 using Least Squared Mean analysis. Plant lines were placed into disease category 1 if their reaction to Sclerotinia blight was not significantly different than the parent genotype Okrun. Alternatively, plant lines were placed into disease category 2 if they demonstrated a significant increase in resistance to Sclerotinia blight when compared to Okrun.
Plant lines were assessed for growth habit type, either spreading (S) or upright (U), as discussed by Smith and Simpson (1995). Any other abnormalities in phenotype, such as stunting, were also noted as part of the plant line growth habit. Plants were harvested individually (October 15 2000–2002) and returned to the laboratory for analysis. Pods (pod mass) were pulled from individual peanut plants and weighed to the nearest gram. Peanuts from individual plant samples were then weighed, counted, and processed through a 16/64 grading screen. Shelling percentage for each sample was calculated by dividing the total kernel weight by the pod mass. Kernels retained on the screen were then weighed and counted. Weight of kernels retained on the screen was calculated by dividing the kernel weight by the kernel count. Statistical analysis was performed using Least Squared Means analysis and Dunnett's test via SAS version 8.2 (Cary, NC).
Results
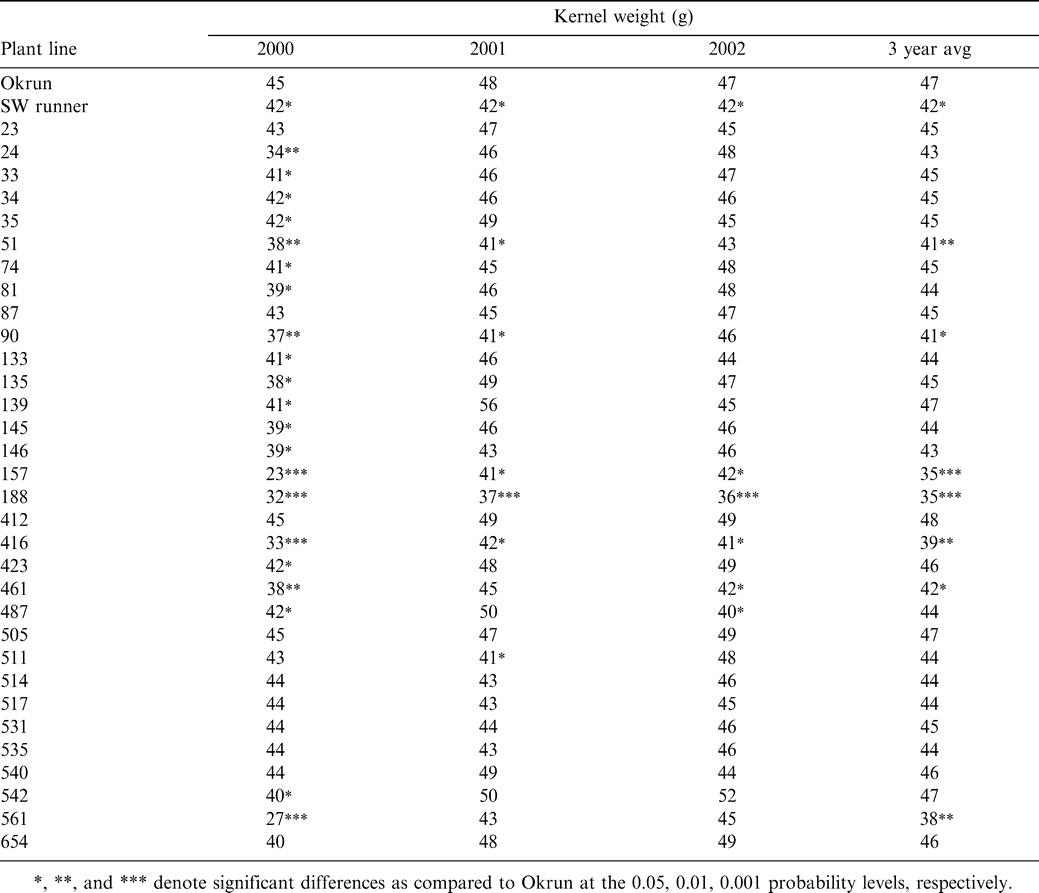
Classification of plant lines into disease and growth habit categories is listed in Table 1. Most plant lines were grouped into disease category 1, demonstrating no significant difference in reaction to Sclerotinia blight than the cultivar Okrun. Fourteen plant lines were placed in disease category 2 over the 3-year period, consistently demonstrating a significant increase in resistance to Sclerotinia blight infection as compared to Okrun. While the majority of plant lines tested did not differ from Okrun in their growth habit, 2 lines had an upright growth habit, with a prominent vertical main stem similar to that of Southwest Runner, and 1 line was noticeably stunted.
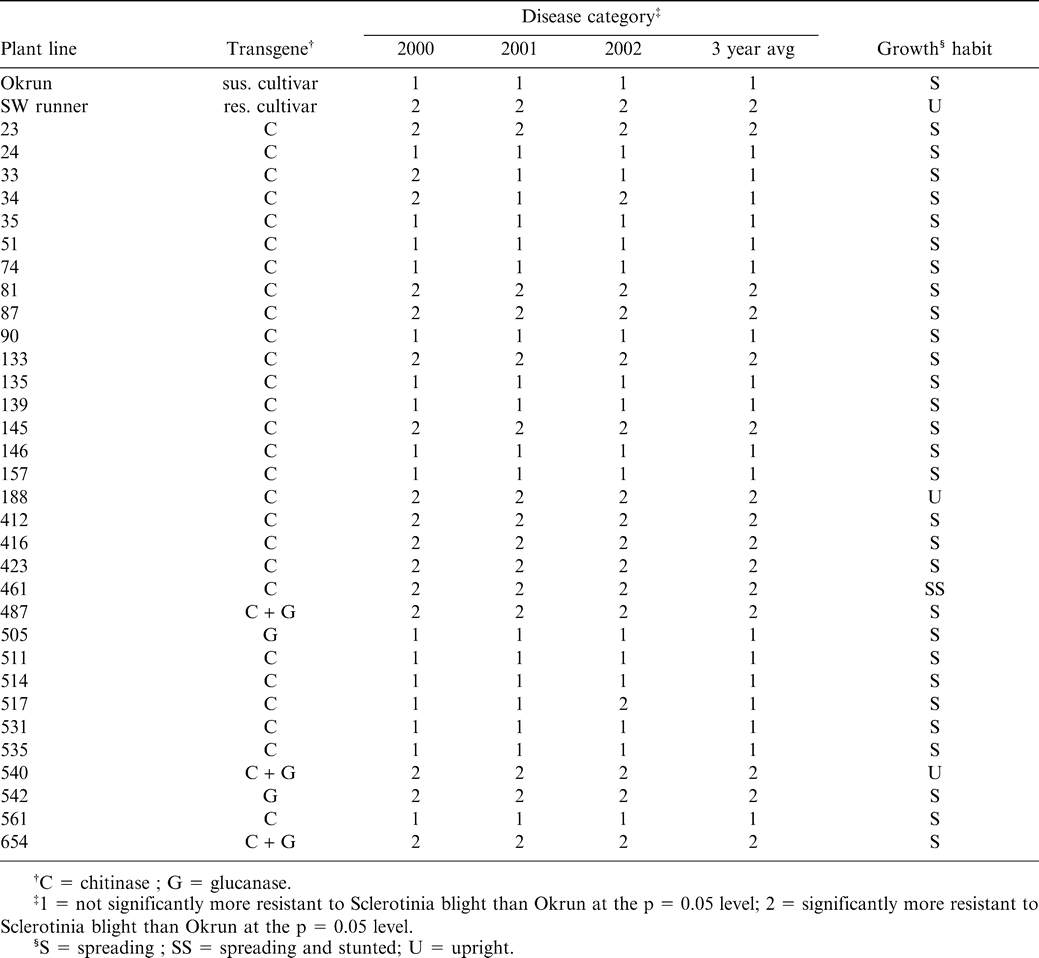
The majority of transgenic test lines did not differ from the parent genotype Okrun with respect to pod mass throughout the 3-year test period (Table 2). In 2000, of the 32 lines tested, 8 lines had a significantly smaller pod mass as compared to Okrun. All lines with reduced pod mass contained only the chitinase transgene, with the exception of line #654 which contains both the chitinase and glucanase transgenes. Reduced pod mass was not always consistent with increased Sclerotinia blight incidence. For example, line #188 never became diseased, but consistently had a lower pod mass than cultivar Okrun. Similarly, pod mass reduction could not be correlated with transgene type due to the fact that the majority of test lines contain only the chitinase transgene and show no variation with respect to Okrun when considering pod mass.
Similar results were recorded in 2001 with the majority of test lines being indistinguishable from Okrun with respect to pod mass (Table 2). Six of the plant lines with reduced pod mass in 2000 continued to have lower pod masses than Okrun in 2001. All plant lines with lowered pod mass contained only the chitinase transgene. Again, pod mass among test lines varied independently of disease incidence and transgene type.
Results from 2002 were less consistent than the two previous years, with only 4 test lines demonstrating a reduced pod mass compared to Okrun (Table 2). These lines (#51, #188, #416, #461) also had a lower pod mass value for other years tested. Over the 3-year period, 8 lines (#51, #74, #157, #188, #461, #561, and #654) averaged a significantly lower pod mass than Okrun.
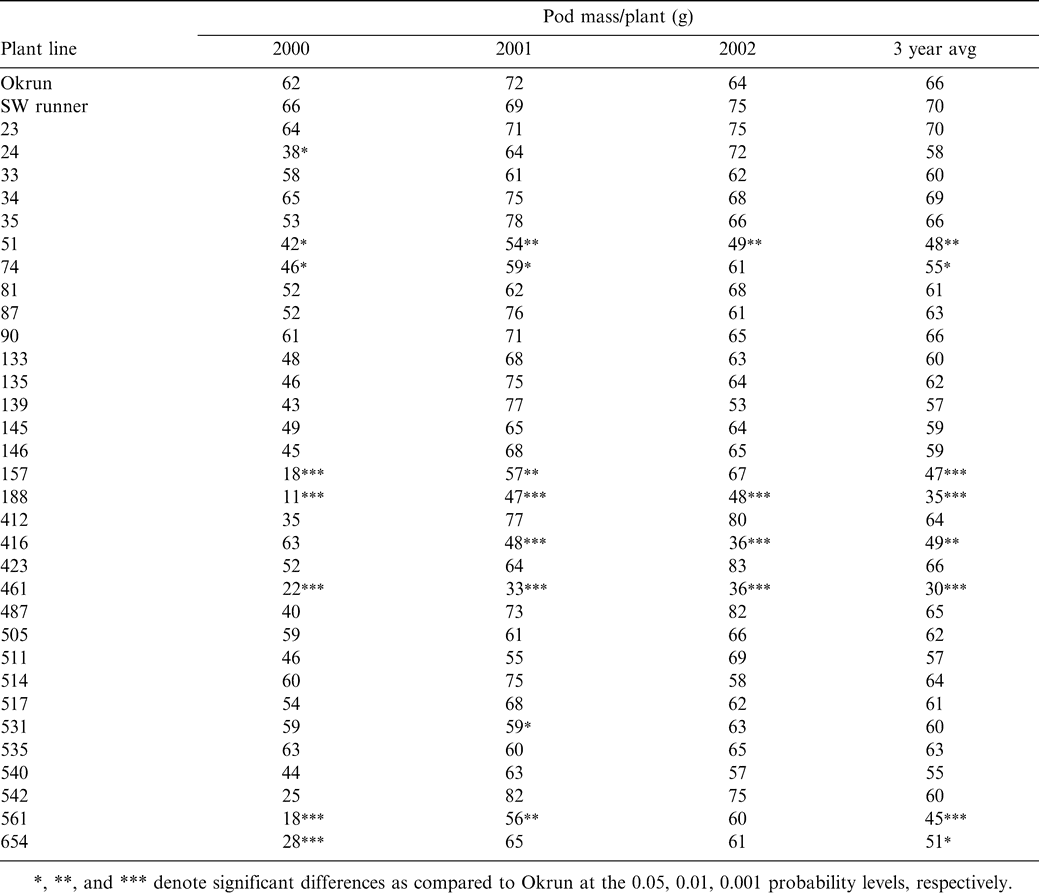
Shelling percentages for all plant lines tested are shown in Table 3. Throughout the 3-year test period, very little deviation from the values reported for the parent genotype Okrun was seen among the transgenic lines tested. The only variation seen was reported in the year 2000, where 3 of the test lines (#157, #188, and #561) had a significantly lower shelling percentage than Okrun. As with pod mass, shelling percentages varied independently of Sclerotinia blight incidence for all lines tested.
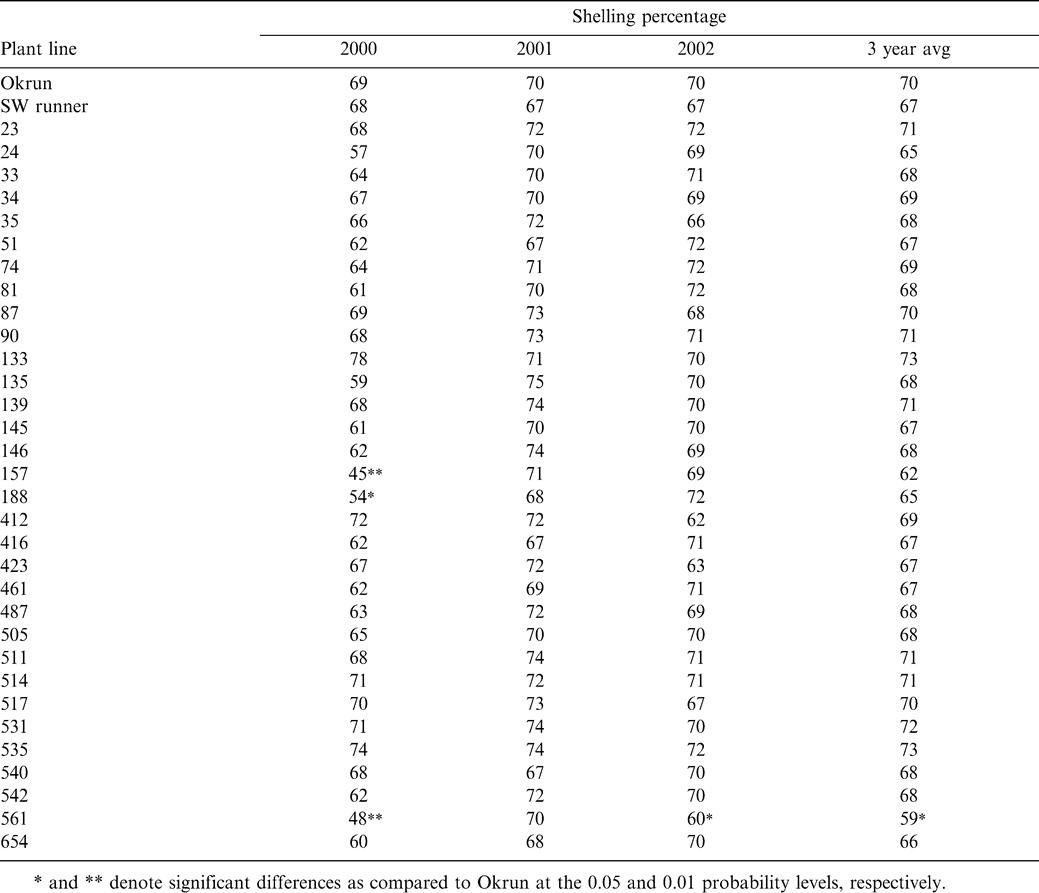
Average kernel weight among transgenic test lines, shown in Table 4, deviated extensively from that recorded for Okrun in the year 2000, but this variation decreased over the 3-year test period. None of the test lines had a kernel weight greater than Okrun. However, 21 lines averaged a kernel weight less than Okrun with the mean decrease being 17%. The number of test lines with kernels significantly smaller than Okrun dropped in 2001 from 21 to six, and then again in 2002 to five. Three of the transgenic lines (#157, #188, and #416) averaged a reduced kernel weight compared to Okrun for each of the three years tested, with the mean decrease being 23%. Average kernel weight was not consistent with Sclerotinia blight incidence. However, there was a slight correlation of average kernel weight and pod mass, with 5 of the test lines (#51, #157, #188, #461, and #561) having a reduction in both quality traits compared to the parent genotype Okrun over the 3-year test period.
Discussion
This study examined the agronomic performance of transgenic peanut plants under natural field conditions to determine the effect that genetic engineering and the introduction of foreign genes may or may not have on agronomic traits and/or yield components. The performance of transgenic plants may be affected by several aspects of the transformation process including, but not limited to: (1) Insertion mutagenesis: insertion of the transgene disrupts the genomic DNA causing an obvious change in phenotype or field performance (Feldmann and Marks, 1987; Feldmann et al., 1989), (2) Pleiotrophy: individual genes, apparently unrelated, affect plant phenotype and performance (Dale and McPartlan, 1992), and (3) Somoclonal variation: genetic variation resulting from the tissue culture and regeneration process itself (Larkin and Scowcroft, 1981; Karp and Bright, 1985; Karp 1991). Somoclonal variation and insertion/expression of the anti-fungal genes caused no differences in the agronomic characteristics and yield components of the majority of the transgenic lines tested as compared to non-transformed Okrun. The largest deviation from non-transformed values occurred in the first year of testing which was probably due to the reduced field fitness of greenhouse grown kernels used for seed. This type of result has been reported previously in field tests of other transgenic crops (Arnoldo and Baszczynski, 1991).
The peanut transformation process did result in consistently deteriorated agronomic traits among 5 of the transgenic lines tested: #51, #157, #188, #461, and #561. Although all of these lines contain the rice chitinase transgene, their decrease in field performance appears to be independent of transgene type since many of the unaffected plant lines also contain the same gene. All affected transgenic lines have a spreading growth habit which upon visible inspection is no different than that of Okrun, with the exceptions of line #188 which grows upright and line #461 which is stunted. It was not determined which aspect of the transformation process caused the phenotypic and agronomic abnormalities among the 5 affected lines.
All of the transgenic peanut lines tested here share the common genetic background of the cultivar Okrun, which is susceptible to fungal infection but when unimpeded, produces peanuts with highly marketable agronomic characteristics. The ultimate goal of this research project is to produce a transgenic peanut line, which retains the desirable agronomic traits of Okrun but has an increased resistance to fungal disease. Several of the peanut lines examined in this study have increased disease resistance and desirable kernel characteristics that warrant further investigation and consideration for commercial release. For example, lines #487 and #540 averaged 50% and 76% less disease, respectively, without loosing any of the yield components characteristic of the Okrun genotype. Assuming 0.91 m row spacing, the average yield for Okrun over the 3-year test period (66 g/plant) can be translated as 3.2 t/ha, with the transgenic plant lines averaging 1.5 to 3.3 t/ha.
Transgenic lines, which have deteriorated agronomic traits may still be of some use in traditional breeding programs. For example, line #188 never became diseased throughout the entire 3-year test period but the kernels from this line are reduced in weight, similar to that of Southwest Runner, which produces kernels of a size undesirable to some shelling companies. Previous studies have shown that morphological resistance exists among lines with upright, bunch growth habits, probably due to the lack of a dense plant canopy which contributes to optimal disease conditions (Melouk and Backman, 1995). The location of transgene insertion for #188 has not yet been determined and thus it is assumed, but unknown, that the insertion event disrupted a gene crucial for a spreading growth habit (mutational interference). Line #188 contains a single copy of the rice chitinase transgene and has been previously shown to have a transgene expression level 22% above background level under non-challenged conditions (Chenault et al., 2002). Although the transgene expression level of line #188 is well within the range of hydrolase activity reported elsewhere for plant varieties with heightened fungal resistance (Lozovaya et al., 1998), we realize the upright growth habit most likely plays a major role in the total resistance observed. Transfer of the resistance trait of line #188 into other genetic backgrounds could eliminate the problem of small kernel size.
Agronomic characteristics of the peanut lines evaluated under field conditions in this study, including pod mass, shelling percentage, and kernel weight, were statistically comparable between the majority of transformed and non-transformed plant lines. These results indicate that peanut can be genetically engineered successfully, and that in this case, plant lines can be regenerated in which the microprojectile bombardment transformation system does not introduce any adverse effects on the intrinsic agronomic and qualitative traits critical to the peanut industry.
Acknowledgements
The authors would like to express their gratitude to the Oklahoma State University Agricultural Experiment Station for their aid in providing land and personnel to conduct these studies. Research assistance provided by Lisa Myers and Doug Glasgow is also deeply appreciated.
Literature Cited
Allefs S. J. H. M. , Florack D. E. A. , Hoogendorn C. , and Stiekeman W. J. 1995 Erwinia soft rot resistance of potato cultivars transformed with a gene construct coding for antimicrobial peptide cecropin B is not altered. Amer. Pot. J 72 : 437 – 445 .
Arce-Ochoa J. P. , Daniello F. , and Pike L. M. 1995 Field performance comparison of two transgenic summer squash hybrids to their parental hybrid line. HortSci 30 : 492 – 493 .
Arnoldo M. and Baszczynski C. L. 1991 Evaluation of transgenic canola plants under field conditions. Genome 35 : 58 – 63 .
Brandle J. E. and Miki B. L. 1993 Agronomic performance of sulfonylurea-resistant transgenic flue-cured tobacco grown under field conditions. Crop Sci 33 : 847 – 852 .
Casas A. M. , Kononowicz A. K. , Zehr U. B. , Tomes D. T. , Axtell J. D. , Butler L. G. , Bressan R. A. , and Hasegawa P. M. 1993 Transgenic sorghum plants via microprojectile bombardment. Proc. Nat. Acad. Sci 90 : 11212 – 11216 .
Chenault K. D. , Burns J. A. , Melouk H. A. , and Payton M. E. 2002 Hydrolase activity in transgenic peanut. Peanut Science 29 : 89 – 95 .
Chenault K. D. , Payton M. E. , and Melouk H. A. 2003 Greenhouse testing of transgenic peanut for resistance to Sclerotinia minor. Peanut Science 30 : 116 – 120 .
Coulombe B. A. , Panter D. M. , Stanton J. J. , and Ward R. G. 1994 Yield performance of transgenic cotton strains resistant to Buctril herbicide. Proc. Beltwide Cotton Conf 2 : 651 .
Delannay X. , Bauman T. T. , Beighley D. H. , Buettner M. J. , Coble H. D. , DeFelice M. S. , Derting C. W. , Deidrick T. J. , Griffin J. L. , Hagood E. S. , Hancock F. G. , Hart S. E. , LaVallee B. J. , Loux M. M. , Lueschen W. E. , Matson K. W. , Moots C. K. , Murdock E. , Nickell A. D. , Owen M. D. K. , Paschal E. H. , Prochasks L. M. , Raymond P. J. , Reynolds D. B. , Rhodes W. K. , Roeth F. W. , Sprankle P. L. , Taroshione L. J. , Tinius C. N. , Walker R. H. , Wax L. M. , Weigelt H. D. , and Padgette S. R. 1995 Yield evaluation of a glyphosate-tolerant soybean line after treatment with glyphosate. Crop Sci 35 : 1461 – 1467 .
D'Halluin K. , Botterman J. , and deGreef W. 1990 Engineering of herbicide-resistant alfalfa and evaluation under field conditions. Crop Sci 30 : 866 – 871 .
D'Halluin K. , Bossut M. , Bonne E. , Mazur B. , Leemans J. , and Botterman J. 1992 Transformation of sugarbeet (Beta vulgaris L.) and evaluation of herbicide resistance in transgenic plants. Bio/tech 10 : 309 – 314 .
Dale P. J. and McPartlan H. C. 1992 Field performance of transgenic potato plants compared with controls regenerated from tuber discs and shoot cuttings. Theor. Appl. Gen 84 : 585 – 591 .
Feldmann K. A. and Marks M. D. 1987 Agrobacterium-mediated transformation of germinating seeds of Arabidopsis thaliana, a non-tissue culture approach. Mol. Gen. Genet 208 : 1 – 9 .
Feldmann K. A. , Marks M. D. , Christianson M. L. , and Quatrano R. S. 1989 A dwarf mutant of Arabidopsis generated by T-DNA insertion mutagenesis. Science 243-1351 – 1354 .
Fuchs M. and Provvidenti R. 1996 Evaluation of transgenic tomato plants expressing the coat protein gene of cucumber mosaic virus strain WL under field conditions. Plant Dis 80 : 270 – 275 .
Karp A. 1991 On the current understanding of somoclonal variation. Oxford Surveys Plant Mol. Cell Biol 7 : 1 – 58 .
Karp A. and Bright S. W. J. 1985 On the causes and origins of somoclonal variation. Oxford Surveys Plant Mol. Biol 2 : 199 – 234 .
Kirby J. S. , Melouk H. A. , Stevens T. E. , Banks D. J. , Sholar J. R. , Damicone J. P. , and Jackson K. E. 1998 Registration of ‘Southwest Runner’ peanut. Crop Sci 38 : 545 – 546 .
Larkin P. J. and Scowcroft W. R. 1981 Somoclonal variation- a novel source of variation from cell cultures for plant improvement. Theor. Appl. Gen 60 : 197 – 214 .
Lin W. , Anuratha C. S. , Datta K. , Potrykus I. , Muthukrishnan S. , and Datta S. K. 1995 Genetic engineering of rice for resistance to sheath blight. Bio/tech 13 : 686 – 691 .
Lozovaya V. V. , Waranyuwat A. , and Widholm J. M. 1998 β-1,3 Glucanase and resistance to Aspergillus flavus infection in maize. Crop Sci 38 : 1255 – 1260 .
Melouk H. A. and Backman P. A. 1995 Management of soilborne fungal pathogens. 75 – 85 In Melouk H. A. and Shokes F. M. eds. Peanut Health Management APS Press St. Paul, MN .
Porter D. M. and Steele J. L. 1983 Quantitative assay by elutriation of peanut field soil for sclerotia of Sclerotinia minor. Phytopath 73 : 636 – 640 .
SAS Inst 1985 SAS/STAT Guide for personal computers, Ver.6 Ed SAS Int Cary, N.C .
Simpson C. E. , Smith O. D. , and Melouk H. A. 2000 Registration of ‘Tamrun 98’ peanut. Crop Sci 40 : 859 .
Smith O. D. , Simpson C. E. , Grichar W. J. , and Melouk H. A. 1991 Registration of ‘Tamspan 90’ peanut. Crop Sci 31 : 1711 .
Smith O. D. and Simpson C. E. 1995 Selection of peanut cultivars. 19 – 22 In Melouk H. A. and Shokes F. M. eds. Peanut Health Management APS Press St. Paul, MN .
Stahl E. A. and Bishop J. G. 2000 Plant-pathogen arms races at the molecular level. Current Opinion in Plant Biology 2000 3 : 299 – 304 .
Tricoli D. M. , Carney K. J. , Russell P. F. , McMaster J. R. , Groff D. W. , Hadden K. C. , Himmel P. T. , Hubbard J. P. , Boeshore M. L. , and Quemada H. D. 1995 Field evaluation of transgenic squash containing single or multiple virus coat protein gene constructs for resistance to cucumber mosaic virus, watermelon mosaic virus2, and zucchini yellow mosaic virus. Bio/tech 13 : 1458 – 1465 .
Vasil V. , Castillo A. M. , Fromm M. E. , and Vasil I. K. 1992 Herbicide resistant fertile transgenic wheat obtained by microprojectile bombardment of regenerable embryonic callus. Bio/tech 10 : 662 – 674 .
Wilson F. D. , Flint H. M. , Deaton W. R. , and Buehler R. E. 1994 Yield, yield components, and fiber properties of insect-resistant cotton lines containing a Bacillus thuringiensis toxin gene. Crop Sci 34 : 38 – 41 .
Yang H. , Singsit C. , Wang A. , Gonsalves D. , and Ozais-Akins P. 1998 Transgenic peanut plants containing a nucleocapsid protein gene of tomato spotted wilt virus show divergent levels of gene expression. Plant Cell Rep 17 : 693 – 699 .
Notes
Author Affiliations
*Corresponding Author: K.D. Chenault, USDA-ARS Wheat, Peanut, and Other Field Crops Research Unit, Stillwater, OK 74075; kelly.chenault@ars.usda.gov