

Introduction
One of the most effective methods of evaluating plant water use is by measuring water flow through the xylem of a plant. This technique, known as sap or stem flow measurement, can continuously monitor water flow without disturbing normal plant functions (Senock et al., 1996), and has been shown to give highly accurate measurements of plant water use (Lascano et al., 1992, Devitt et al., 1993, Dugas et al., 1993). Despite the use of sap flow measurement in several different crops (Dugas et al., 1994; Senock et al., 1996; Gordon et al., 1999; Lascano, 2000; Lee et al., 2002; Li et al., 2002; Lu et al., 2002), very few studies have directly examined water flow in peanut. Erickson et al. (1986) used the heat pulse method to measure apparent sap flow velocities in peanut genotypes but this was done in a greenhouse for very limited time periods (1100 to 1300 hour apparent solar time).
The quantification of stem water flow could provide very critical information towards solving a wide variety of problems in peanut production. These include: quantifying the effects of insect damage to plant function; determining how changes in peanut water use may affect pre-harvest aflatoxin production; or evaluating the efficacy of irrigation scheduling or irrigation methods in peanut production. To determine the actual differences in plant water-use among irrigation methods for peanut, it is important to quantify plant water use on a daily basis and under typical field conditions. Sap flow measurements provide a means to accomplish this. However, the lack of verification of this technique in peanut remains a barrier to the utilization of this technology.
Maintaining efficiency in irrigation remains a major problem in almost every U.S. peanut producing region. Irrigated peanut acreage comprises over 50% of all peanut production in the U.S. and can increase yields by up to 19% over non-irrigated production (Lamb et al., 1997). But in utilizing irrigation, it is important to maximize agronomic water-use efficiency while maintaining optimum production. The primary method of irrigation in peanut production is overhead sprinkler irrigation (OH) (Anon., 1996). An alternative irrigation method that promises to provide greater agronomic water-use efficiency over typical center pivot or lateral sprinkler systems is subsurface drip irrigation (SDI) (Phene et al., 1992; Sorensen et al., 2001; Assouline et al., 2002). However, despite the purported benefits of SDI over OH irrigation, few studies have quantified the actual differences in plant water use between the two systems.
For this study, peanut sap flow measurements were taken to evaluate the efficacy of the technique under field conditions and to determine if the technique could detect differences in plant water use between irrigation methods. In addition, if differences in peanut water use existed, crop biomass production and carbon isotope discrimination were measured to determine the differences in water allocation strategies between plants in different irrigation schemes. The specific objectives of this research were to: 1) determine if the technique of sap flow measurement could be successfully utilized in peanut; 2) determine if daily peanut stem water flow, biomass partitioning, and carbon isotope discrimination differed among irrigation treatments, and 3) determine what environmental parameters were most closely correlated with peanut stem water flow.
Materials and Methods
Field Conditions
The experiment was conducted during the 2001 crop year at the USDA-ARS Multi-crop Research Farm in Shellman, Georgia. The soil was a Greenville fine sandy loam (fine, kaolinitic, thermic Rhodic Kandiudults) with 0–2% slope. Conventional tillage practices were followed and included: disking, subsoiling, moldboard plowing, field cultivating, rototilling, and planting. Three side by side experimental fields were established at this research farm, one ca. 146 m × 329 m with overhead (OH) irrigation treatments, one ca. 98 m × 329 m with subsurface drip irrigation (SDI) treatments, and one ca. 49 m × 329 m, located between the overhead and subsurface drip fields serving as the non-irrigated (NI) treatment. Each field was divided into 16.5 m × 36.6 m plots containing 18 cropping rows and 2 border rows each containing crop rotations and replications.
The lateral overhead irrigation system contained three equal length spans with approximately 13 nozzles each. To apply differential levels of water to each treatment plot, different sized sprinkler heads were placed on each drop nozzle within a span, such that one span applied a full rate of desired irrigation (100%), and the second span applied 66% of this amount. Overhead sprinkler irrigation amount and timing were determined by the Irrigator Pro expert system (Lamb et al., 1993; Davidson et al., 1998) based on soil temperature measurements collected in the 100% treatment. Time between irrigation events was typically three to four days during the most active crop growth period of the season.
The SDI system consisted of drip tube laterals spaced 0.91 m apart underneath each crop row at a depth of 0.25–0.3 m. The drip tubing (NetafimTM, Tel Aviv, Israel) was 0.38 mm thick (15 mil) with emitters spaced at 45 cm and a flow rate of 1.48 liters hour−1 emitter−1 for the 100% treatment, and 0.91 liters hour−1 emitter−1 for the 62% treatment. An on-site weather station collected meteorological data and estimated potential evapotranspiration (ETo) using the modified Jensen-Haise equation corrected for local conditions. Irrigations were scheduled daily with plant evaporated water being replaced using crop coefficients (Harrison and Tyson, 1993). Estimated ETo was multiplied by the crop coefficient and rainfall was subtracted to estimate actual evapotranspiration (ETa). If rainfall was greater than ETa no irrigation was applied.
The Georgia Green peanut cultivar was planted in OH, SDI, and NI plots on 24 May 2001 in twin rows spaced 0.18 m apart using a Monosem 8812 twin row vacuum planter (ATI Inc., Lexena, KS). Eight seeds ft−1 (26.2 seeds m−1) were planted in twin rows centered at 0.91 m. Approximately 60 days after planting, sensors and associated dataloggers were installed in the fields to measure sap flow, soil moisture, canopy surface temperature, and soil temperature. On 5 October 2001 all treatments were mechanically harvested and samples were collected for measurements of yield and grade.
Sap Flow Measurements
Two Flow32 Sap Flow Systems (Dynamax, Inc.) with a total of 16 Microsensors (SGA5) were installed to measure sap flow. Because of the limitation of cable length with the sap flow system, only one 16.5 m × 36.6 m plot of each irrigation type (100% OH, 66% OH, 100% SDI, 62% SDI, and NI) could be measured. Sap flow gauges were installed on peanut stems 11 September 2001 at a phenological stage of high physiological activity and active pod filling (Ketring et al., 1982) and where stems had reached maximum size to facilitate installation of the gauges. Sap flow gauges were installed on randomly located plants within each irrigation treatment with one gauge per plant for a total of: four gauges in the 100% OH plot, four gauges in the 66% OH plot, three in the NI plot, two in the 62% SDI plot, and three in the 100% SDI plot. Gauges were installed on peanut main stems, standardizing the position at approximately 10–15 cm above the ground and where stem internodes were long enough to permit a gauge. Stem diameters of the internodes on which the gauges were placed ranged from 0.3 to 0.5 cm. On 25 September 2001, all gauges were removed and reinstalled on different plants. The stem diameters of the internodes where gauges were placed ranged from 0.3 to 0.45 cm and the number of leaves ranged from 5 to 9 above the gauge (at time of installation). Sap water flow data was collected at 15 minute intervals on a 24 hour basis. During the period between 15 September–29 September 2001 (258–272 day of year), the most consistent daily data was collected from the majority of the gauges. However, usable data was only available for a single gauge in the 62% SDI treatment and is not reported. For consistency among measurements, data from two gauges in the 100% OH, two gauges in the 100% SDI, and three gauges in the NI treatments were used for water use comparisons.
Environmental and Canopy Monitoring
Environmental sensors and associated dataloggers were installed (simultaneous to sap flow gauge installation) to record leaf and soil temperatures and soil moisture on an hourly basis. In the same row and within 60 cm of each plant with a sap flow gauge, hourly data was recorded measuring: 1) canopy surface leaf temperature using infrared thermometers (model IRt/c.2, Exergen, Watertown, MA); 2) soil moisture (ECH2O, Dielectric Aquamater, Decagon Devices, Inc., Pullman, WA) at two ranges: shallow (5–25 cm) and deep (30–50 cm); and 3) soil temperature at 5 cm depth using thermocouples.
Carbon Isotope Analysis
In order to determine seasonal plant water-use efficiency (WUE), peanut leaf tissue was collected from 6 plants (one leaf each) located near sap flow gauges within each irrigation treatment approximately 90 days after planting on 24 August 2001 and used to measure carbon isotope discrimination. This sampling date corresponds with the phenological period associated with the highest rubisco levels and, concomitantly, the highest photosynthetic levels of the season. Differences in photosynthesis (and therefore WUE) due to irrigation would be most evident at this time period (Nageswara Rao and Wright, 1994; Nageswara Rao et al., 1995). Tissue collection was standardized to second nodal apex leaves with minimal or no insect or disease damage. Standardizing to the second nodal leaf position has been shown to maximize the relationship between chlorophyll content and specific leaf area (Nageswara Rao et al. 2001). Tetrafoliate leaves were excised and chlorophyll content was measured on these leaves in the field using the Minolta SPAD chlorophyll meter (Minolta Corp., Ramsey, N.J.). The SPAD chlorophyll meter measures plant tissue absorbance of wavelengths in the visible spectrum and serves as a measure of the relative internal concentration of chlorophylls a and b. One SPAD chlorophyll reading was taken on each of the four leaflets, avoiding the midrib, and then averaged for one chlorophyll reading per plant to correct for possible non-homogeneous distribution of chlorophyll throughout the leaf (Monje and Bugbee 1992). Tetrafoliate leaves were then placed on ice and refrigerated at 4 C until further analysis.
In the laboratory, the field collected leaves were hydrated in distilled water for at least three hours prior to leaf area measurements. Four leaflets were removed from each petiole and their areas measured with an LI-3000A leaf area meter (LI-COR Inc., Lincoln, NE), and summed to give total leaf area. Leaflets were then oven dried at 60 C for 72 hours and weighed. Specific leaf area (SLA) was calculated as the ratio of leaf area to leaf dry weight. Leaflets were then fine ground using a Braun ® (model KSM2) coffee grinder and analyzed for carbon isotope discrimination ratio (δ13C).
The isotopic discrimination in the peanut leaf samples was analyzed for δ13C at the University of Arkansas Stable Isotope Laboratory. Samples of the ground leaves (2 mg, +/− 0.2 mg) were weighed, sealed in capsules and, along with standards, loaded into an elemental analyzer autosampler (a “Zero Blank” autosampler from Costech Analytical Technologies in Valencia, CA). Samples and standards were then combusted in the elemental analyzer (Carlo Erba NC2500 elemental analyzer coupled with a Thermoquest Finnigan Delta plus isotope ratio mass spectrometer). Lab standards, which were calibrated against internationally distributed isotope standards, were analyzed at regular intervals throughout the sample runs. The resulting N2 and CO2 gases (along with isotopic reference gases for N2 and CO2) were admitted to the mass spectrometer via Finnigan's Conflo II interface. Data were collected and processed by Finnigan's Isodat software. Sample results were based on one analysis per sample (δ15N, δ13C, %N and %C were all determined with the same analysis). Isotope results were reported in delta notation vs. air (for nitrogen) and vs. PDB (for carbon) in permil. Stable carbon isotope composition was expressed as δ13C, where δ13C (‰) = [(R sample/R standard)−1] × 1000, and R is the 13C/12C ratio.
Biomass Measurements
Samples of both apex stem tissue and whole plants were collected from the same plots containing the sap flow gauges and measured for leaf and stem partitioning. When sap flow collars were changed on 25 September 2001 and removed for the final harvest on 3 October 2001, those stems with attached gauges were collected from all irrigation treatments and divided into leaves and internodes. Leaves were then hydrated and leaf area measured as with the isotope samples. Internode length was measured to the nearest mm, and leaf and internode tissue dry mass determined. On 3 October 2001, four whole plants in each irrigation treatment were collected by cutting the aboveground portion of the plant at the soil level. Whole-plant tissue was separated into stems and leaves, and whole plant leaf area, leaf dry mass, and stem dry mass were measured. Specific leaf area was also calculated. Yield in dry mass of pods (standardized to 7% moisture) from each plot were recorded at harvest.
Leaf Area Index
Leaf area index (LAI) was measured on 1 October 2001 with a Li-Cor LAI-2000 Plant Canopy Analyzer. Measurements were taken at six locations within each irrigation treatment that contained sap flow gauges. At each location and using a 90 degree angle cap, one measurement was taken above the canopy and four below the canopy. Two repetitions were used, one with the wand pointed down the row and the other with the wand pointed perpendicular to the row.
Statistics
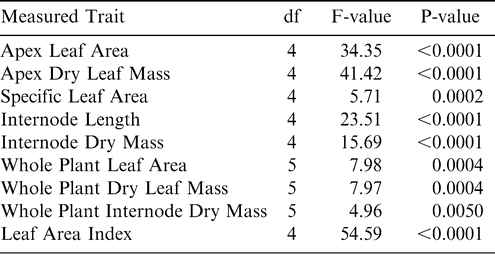
Statistical analyses were performed using SAS and SAS JMP (SAS, 1997). The general linear models (GLM) procedure and analysis of variance (ANOVA) were used to determine differences among irrigation treatments for: average daily water use; cumulative water use over a 24 hour period; biomass characteristics of apex stems and total plants; leaf area index; and δ13C, SLA, and SPAD chlorophyll measurements on the same leaf tissue. Apex internode dry mass was log transformed to conform to the normality assumptions of ANOVA. Pearson correlations were used to evaluate possible relationships of hourly sap flow rates and environmental characteristics.
Results
Sap flow was successfully measured in peanut during a two week period in all irrigation treatments. Figure 1 illustrates the daily fluctuations of water use measured in the three irrigation treatments and their responses to irrigation and/or rainfall. Peanut water use characteristically peaked during midday with no flow or very low levels of water flow at night. Typical daily maximum flow rates ranged from 4–6 g/hr in irrigated treatments and from 1–2 g/hr in the NI treatment. Decreased water use in response to rainfall was recorded in all treatments during the day the water was received. However, in SDI plots, water use apparently was not dampened by the daily application of subsurface water (Fig. 1). The sap flow technique was also able to record minute changes in peanut water flow during the day presumably in response to changing edaphic and climatic factors (Fig. 1).
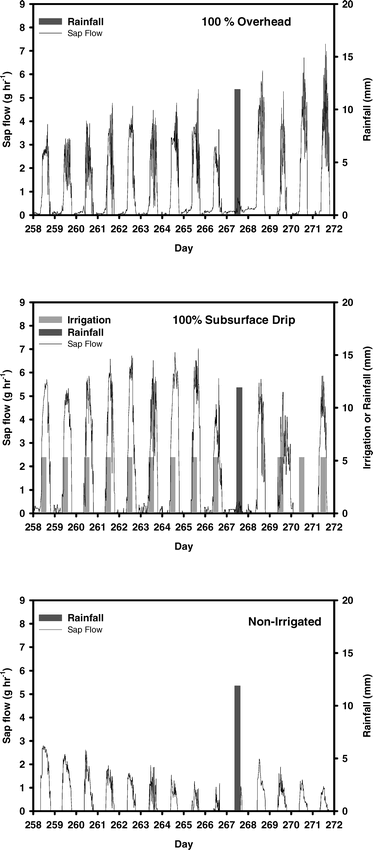
Daily sap flow of Georgia Green peanuts under full level of irrigation in overhead (100% overhead irrigation) and subsurface drip irrigation (100% subsurface drip), and in non-irrigated treatments. Plots show sap flow (grams of water per hour) by day for the period day 258 (15 Sept. 2001) through 272 (29 Sept. 2001). Superimposed on sap flow are a single precipitation event and numerous irrigation events for subsurface drip. No irrigation was applied in the OH treatment during this time period.
Significant differences in peanut water use as measured by sap flow gauges existed among NI, SDI, and OH treatments. Statistical analysis of two replications each in OH and SDI treatments and three replications in the NI treatment showed significant differences among irrigation treatments in both average daily sap flow rate (df = 2, F = 32.66, P-value = <0.0001; R-square = 0.76) and cumulative sap flow over a 24 hour period (df = 2, F = 32.19, P-value = <0.0001; R-square = 0.76). Plants in the SDI treatment had significantly higher average sap flow rates and total water use during a 24 hour period than plants in the OH or NI treatments (Fig. 2).
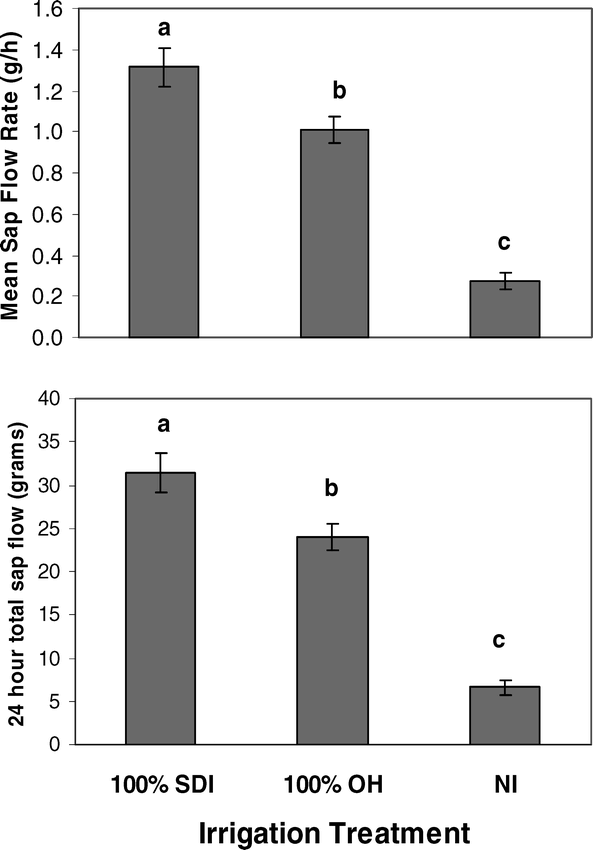
Differences in mean peanut sap flow rate and 24 hour cumulative sap flow among full irrigation levels of subsurface drip (100% SDI) and overhead irrigation (100% OH), and non-irrigated (NI) treatments. Letters denote significant differences among treatments.
Peanut sap flow was significantly correlated on an hourly basis with most of the environmental conditions measured across all irrigation treatments: soil moisture, soil temperature, and infrared canopy temperature (Table 1). However, it was not possible to correlate sap flow with these environmental parameters for the NI treatment due to large gaps in the environmental data for this treatment. When both 100% SDI and 100% OH treatments were pooled, sap flow was significantly positively correlated with: soil moisture at 30–50 cm depth; the average of the shallow and deep soil moisture; soil temperature at the five cm depth; and infrared canopy temperature (Table 1). The most significant relationship was found between infrared canopy temperature and sap flow. Examining irrigation treatments individually revealed that sap flow was significantly correlated with shallow soil moisture, average soil moisture, soil temperature, and infrared canopy temperature in the SDI treatment. In the OH treatment, sap flow was not correlated with soil moisture at either measured depth, but was significantly negatively correlated with soil temperature and positively correlated with infrared canopy temperature (Table 1).
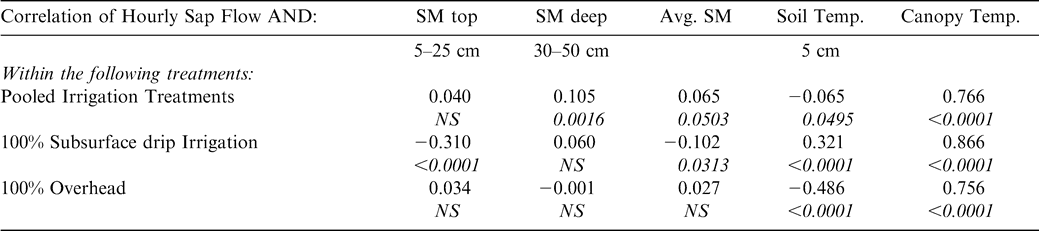
Pearson Correlation Coefficients and P-values between hourly sap flow and: soil moisture (SM) at two depths (top = 5–25 cm, deep = 30–50 cm), the average of the soil moisture readings at these two depths, soil temperature at 5 cm depth, and mean leaf canopy temperature. Correlations run for both irrigation treatments (100% subsurface drip, and 100% overhead) combined (pooled irrigation treatments) and for individual irrigation treatments.
Differences in plant biomass among irrigation treatments revealed that overall plant biomass was greatest in the SDI treatments. There were significant differences among irrigation treatments for apex stem biomass traits, whole plant leaf area and biomass, and leaf area index (Table 2). On apex stems, average leaf area and internode length were significantly greater for both SDI levels than for either level of OH treatment or for the NI treatment (Fig. 3). Specific leaf area was significantly lower for the 100% SDI treatment than for any other treatment (Fig. 3), indicating that leaves in this treatment were significantly thicker. Further, whole plant biomass was generally larger in both levels of the SDI treatments than either level of OH irrigation or the NI treatment (Fig. 4). This included larger total leaf area, total leaf weight and total internode weight, with most comparisons showing significant differences between SDI levels and OH or NI treatments (Fig. 4). This increased aboveground biomass was also evident in significantly larger leaf area indices for both SDI levels than for either OH level or the NI treatment (Fig. 5).
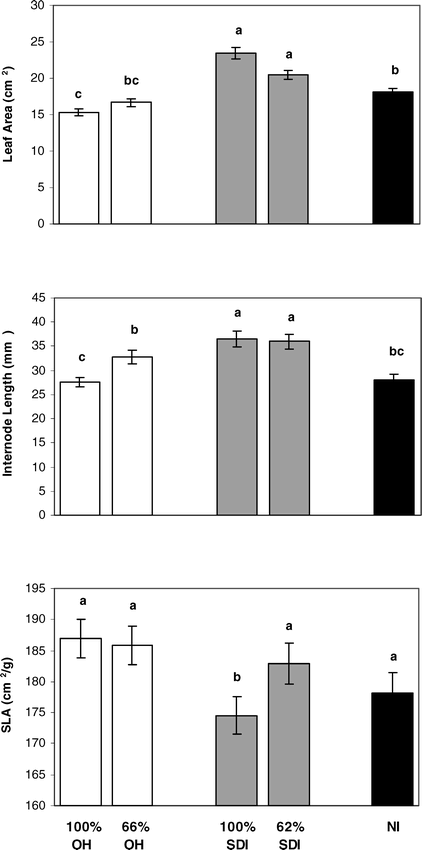
Differences in peanut apex stem biomass traits among: full and partial levels of overhead irrigation (100% OH, 66% OH); full and partial levels of subsurface drip irrigation (100% SDI and 62% SDI); and non-irrigated (NI) treatments. Traits include leaf area, internode length, and specific leaf area (area/mass). Letters denote significant differences among treatments within a trait.
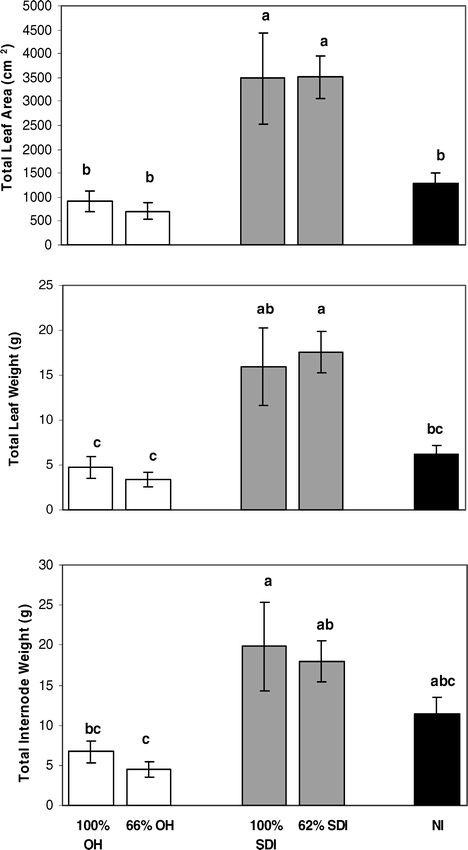
Differences in peanut total plant biomass traits among: full and partial levels of overhead irrigation (100% OH, 66% OH); full and partial levels of subsurface drip irrigation (100% SDI and 62% SDI); and non-irrigated (NI) treatments. Traits include total plant leaf area, total plant leaf weight, and total plant internode weight. Letters denote significant differences among treatments within a trait.
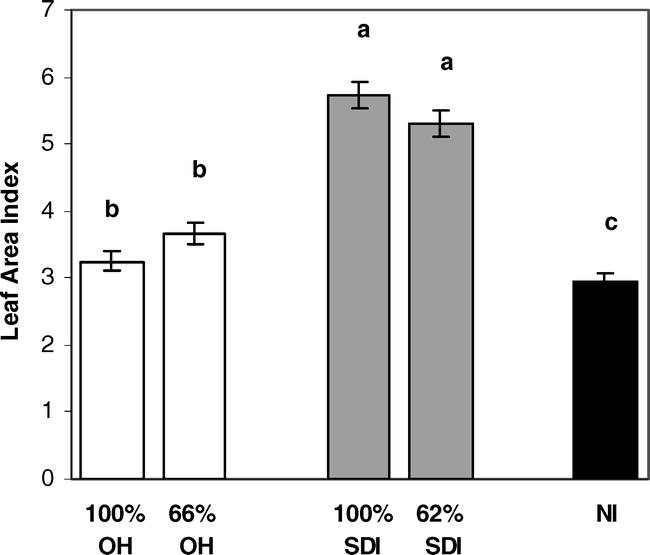
Leaf Area Index differences among: full and partial levels of overhead irrigation (100% OH, 66% OH); full and partial levels of subsurface drip irrigation (100% SDI and 62% SDI); and non-irrigated (NI) treatments. Letters denote significant differences among treatments.
There were no significant differences in seasonal water-use efficiency among irrigation treatments as measured by carbon isotope discrimination at 90 days after planting, although trends were apparent (Fig. 6). Carbon isotope discrimination indicated that plants in both SDI levels tended to have lower water-use efficiencies than either OH levels or the NI treatment, but these differences were not statistically significant. SPAD chlorophyll content (measured on the same leaves that were analyzed for isotope composition) did differ among irrigation treatments with slightly lower chlorophyll levels in the SDI treatments than in either OH or NI treatments (Fig. 6).
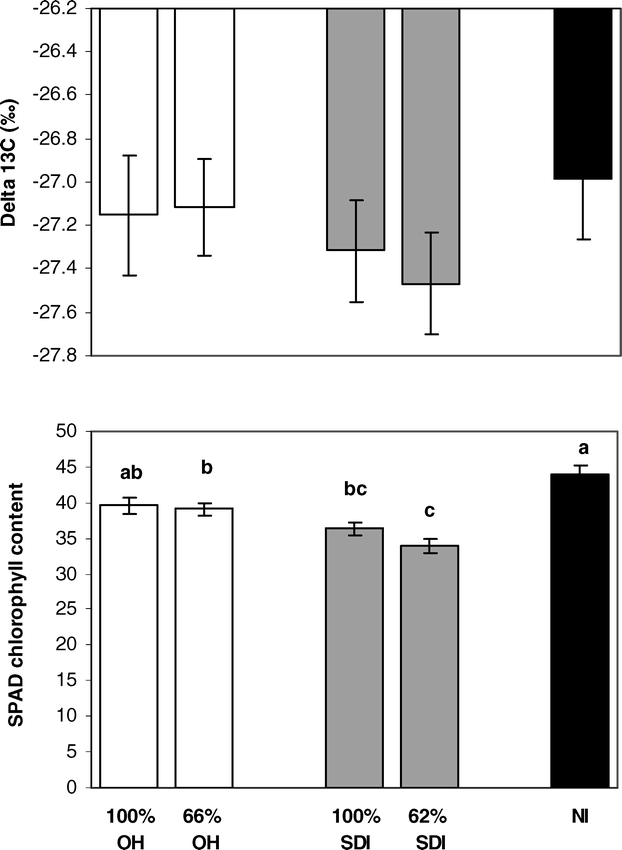
Differences in peanut tissue collected for carbon isotope analysis among: full and partial levels of overhead irrigation (100% OH, 66% OH); full and partial levels of subsurface drip irrigation (100% SDI and 62% SDI); and non-irrigated (NI) treatments. Traits include δ13C and SPAD chlorophyll content measured on the same leaf tissue. Letters denote significant differences among treatments.
Although there was increased water use and increased aboveground biomass in the SDI vs. OH treatments, there was no concomitant increase in pod yield for SDI plants. Yields were not significantly different between OH and SDI treatments when compared across the three replications within the whole field (data not shown). Yields within the plots containing the sap flow collars were: 2180 and 2210 kg for the 100% and 66% OH respectively; 1997 and 2212 kg for the 100% and 62% SDI respectively; and 1441 kg for the NI treatment.
Discussion
This study has demonstrated that sap flow gauges can be used successfully to quantify stem water flow in peanuts in a field production setting. Historically, previous studies have had great success in quantifying sap flow and the effects of many environmental conditions and treatments on water use in: trees (Hatton et al., 1995; Sorensen et al., 1999); shrubs (Allen and Grime, 1995); rangeland plants (Dugas et al., 1992; Senock and Ham, 1995); and crops (Senock et al., 1996; Gordon et al., 1999; Lee et al., 2002; Lu et al., 2002), including corn (Li et al., 2002) and cotton (Dugas et al., 1994; Lascano, 2000). However, only one previous study has examined sap flow in peanuts under greenhouse conditions (Erickson et al., 1986).
The results in this study indicate that peanut plants in the SDI treatment used significantly more water during the late season sampling period than plants in either OH or NI treatments. This was likely due to the larger plant size, as evidenced in both leaf area and internode lengths, of the SDI plants. Because there were no significant differences in yield among irrigated treatments, and given the greater water use of SDI vs. OH plants, the SDI plants in this study appeared to have lower WUE than OH plants. However, this may not be due to the technique of SDI per se but in the management system used in this study to schedule irrigation.
The current irrigation scheduling systems used in both the OH and SDI treatments were based on principles defined as deficit irrigation. Deficit irrigation is also known as the engineering approach and involves irrigating at a level below what the crop demands. Such deficits are generally determined by taking reference evapotranspiration (ETo) and multiplying it by a crop coefficient (Allen et al., 1998; Lascano, 2000). In this study, this was done directly in the SDI treatment using a modified Jensen-Haise equation and indirectly in the OH treatment using soil temperatures and equations that relate crop coefficients to irrigation required in the Irrigator Pro program. For cotton grown on the Texas High Plains, the engineering approach was found to inadequately describe the daily ET of the crop and was less sensitive to the dynamic nature of soil evaporation (Lascano, 2000). In the current study, it appears that the engineering approach also failed to describe the actual water use of peanuts but in the direction of overestimating crop evapotranspiration. Consequently, both irrigation treatments received more water than was needed, especially in the SDI plants. The overestimation of deficits may have been due primarily to outdated crop coefficients which were developed for varieties no longer grown in peanut production (Stansell et al., 1976).
Sap flow in peanuts was significantly correlated with several environmental parameters. Such significant correlations are extremely important for possible applications in irrigation scheduling and crop stress monitoring. Soil temperature and peanut canopy temperature are more easily measured than sap flow, and are often assumed to be indicative of plant available water. Nevertheless, validation of such assumptions are rarely confirmed by direct measurements of plant water use (e.g., Fortin and Moon, 1999; Hochman et al., 2001; Morgan et al., 2003). However, validations are essential before assuming that soil or externally measured plant traits correspond with plant water use. This study has directly correlated actual peanut sap water flow with two soil parameters (moisture, temperature) and with peanut canopy temperature and has illustrated that some traits are better than others as predictive tools for plant water use or drought stress.
The most significant correlations of peanut sap flow were with canopy temperature regardless of irrigation treatment. This is not surprising because leaf or canopy temperature has been used as an indicator of plant water stress under many environments for years (Tanner, 1963; Idso et al. 1981; Jackson et al., 1981; Jones 1999). The significant correlation of sap flow with soil temperature provides supportive evidence for the use of Irrigator Pro, an irrigation scheduling program used extensively in peanut production, which relies primarily on soil temperature as a trigger for irrigation (Lamb et al., 1993; Davidson, et al. 1998). It was surprising that soil moisture was not a good indicator of plant water status in the OH treatment during this experiment. Surface application interface may have been a factor, i.e. a wet surface soil in the OH treatment did not necessarily reflect lower moisture levels of deeper soil layers which may have impeded plant water use. However, surface soil moisture was a good indicator of peanut sap flow in the SDI treatment, and is more likely to indicate soil moisture status as a whole in this treatment since water was applied below the surface and would reach deeper soil depths quickly.
The differences in plant biomass among irrigation treatments indicate that the additional water flow measured in peanut stems in the SDI treatment may be allocated more to aboveground plant biomass than final yield. Plants in the SDI treatment were larger and had more leaf mass, longer internodes, and overall larger canopies than either the OH or NI plants (Figs. 3, 4, 5). Interestingly, for all biomass traits, NI plants were very similar in size and biomass to the OH treatment plants (Fig. 4). This may have been due to NI plants receiving adequate early season precipitation (data not shown) to attain large size in mid-season. However, measured water flow in the OH plants was substantially higher than in the NI plants (Figs. 1 and 2) during the active pod fill stage, thus indicating that these NI plants experienced some late-season drought stress.
The high water-use of the SDI treatment plants without increased yield indicates SDI plants may have had lower water-use efficiency than OH plants in this study. Carbon isotope discrimination, though not statistically significant, indicated that plants in both the 100% and 62% irrigation levels in the SDI tended to exhibit lower WUE than either OH or NI treatments. The inability to detect significant differences may be due to inadequate sample sizes because significant effects of irrigation amount on carbon isotope discrimination have been quantified by using larger sample sizes at this experimental site (data unpublished). However, the apparent lower WUE of SDI plants in comparison to OH plants was most likely due to the effect of irrigation management and not on an inherent characteristic of SDI in general.
This study has successfully demonstrated the technique of sap flow measurement in peanut under field conditions which had not been previously demonstrated to date. The technique was further able to quantify water use differences between irrigation treatments. While this study found a higher plant water use in SDI treatments as compared to OH treatments, further studies need to be conducted before conclusions about differential water use patterns between irrigation treatments can be made. The successful application of the sap flow technique in this study shows the efficacy of applying this technique in future studies addressing this question.
Acknowledgements
We thank Kathy Gray for her excellent aid in design, implementation, and analysis throughout this experiment. We thank Larry Powell and John Gardner for invaluable field and technical assistance. We thank Lori Riles for superb statistical analysis consultation. We dedicate the manuscript to the excellent mentoring, advisement, and overall support of Paul Blankenship.
Literature Cited
Allen S. J. and Grime V. L. 1995 Measurements of transpiration from savannah shrubs using sap flow gauges. Agricultural and Forest Meteorology 75 : 23 – 41 .
Allen R. G. , Pereira L. S. , Raes D. , and Smith M. 1998 Crop Evapotranspiration: Guidelines for computing water requirements. FAO Irrigation and Drainage Paper 56, Rome.
Anon 1996 1995 irrigation survey reflects steady growth. Irrigation Journal Jan./Feb : 24 – 39 .
Assouline S. , Cohen S. , Meerback D. , Harodi T. , and Rosner M. 2002 Microdrip irrigation of field crops: effect on yield, water uptake, and drainage in sweet corn. Soil Soc. Am. J 66 : 228 – 235 .
Davidson J. I. , Griffin W. J. , Lamb M. C. , Williams R. G. , and Sullivan G. 1998 Validation of EXNUT for scheduling peanut irrigation in North Carolina. Peanut Science 25 : 50 – 58 .
Devitt D. A. , Berkowitz M. , Schulte P. J. , and Morris R. L. 1993 Estimating transpiration for three woody ornamental tree species using stem-flow gauges and lysimetry. HortScience 28 : 320 – 322 .
Dugas W. A. , Heuer M. L. , Hunsaker D. , Kimball B. A. , Lewin K. F. , Nagy J. , and Johnson M. 1994 Sap flow measurements of transpiration from cotton grown under ambient and enriched CO2 concentrations. Agricultural and Forest Meteorology 70 : 231 – 245 .
Dugas W. A. , Heuer M. L. , and Mayeux H. S. 1992 Diurnal measurements of honey mesquite transpiration using stem flow gauges. J. Range Management 45 : 99 – 102 .
Dugas W. A. , Wallace J. S. , Allen S. J. , and Roberts J. M. 1993 Heat balance, porometer, and deuterium estimates of transpiration from potted trees. Agric. For. Meterorol 64 : 47 – 62 .
Erickson P. I. , Melouk H. A. , and Ketring D. L. 1986 Apparent sap flow velocity in peanut genotypes susceptible and resistant to Verticillium dahliae. Phytopathology 76 : 500 – 502 .
Fortin M. C. and Moon D. E. 1999 Errors associated with the use of soil survey data for estimating plant-available water at a regional scale. Agronomy Journal 91 : 984 – 990 .
Gordon R. , Brown D. M. , Madani A. , and Dixon M. A. 1999 An assessment of potato sap flow as affected by soil water status, solar radiation and vapour pressure deficit. Can. J. Soil Science 79 : 245 – 253 .
Harrison K. and Tyson A. 1993 Irrigation scheduling methods. Bulletin no. B-974 Ga. Agric. Exp. Stn Athens, GA .
Hatton T. J. , Moore S. J. , and Reece P. H. 1995 Estimating stand transpiration in a Eucalyptus populnea woodland with the heat pulse method: measurement errors and sampling strategies. Tree Physiology 15 : 219 – 227 .
Hochman Z. , Dalgliesh N. P. , and Bell K. L. 2001 Contributions of soil and crop factors to plant available soil water capacity of annual crops on Black and Grey Vertosols. Aust. J. Agric. Res 52 : 955 – 961 .
Idso S. B. , Jackson R. D. , Pinter P. J. , Reginato R. J. , and Hatfield J. L. 1981 Normalizing the stress-degree-day parameter for environmental variability. Agricultural Meteorology 24 : 45 – 55 .
Jackson R. D. , Idso S. B. , Reginato R. J. , and Pinter P. J. 1981 Canopy temperature as a drought stress indicator. Water Resour. Res 17 : 1133 – 1138 .
Jones H. G. 1999 Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agricultural and Forest Meteorology 95 : 139 – 149 .
Ketring D. L. , Brown R. H. , Sullivan G. A. , and Johnson B. B. 1982 Growth physiology. In: Peanut Science and Technology Pattee H. E. and Young C. T. eds. American Peanut Research and Education Society, Inc Yoakum, TX, USA .
Lamb M. C. , Davidson J. I. , and Butts C. L. 1993 Peanut yield decline in the southeast and economically feasible solutions. Peanut Science 20 : 36 – 40 .
Lamb M. C. , Davidson J. I. , Childre J. W. , and Martin N. R. 1997 Comparison of peanut yield, quality, and net returns between nonirrigated and irrigated production. Peanut Science 24 : 97 – 101 .
Lascano R. J. 2000 A general system to measure and calculate daily crop water use. Agron. J 92 : 821 – 832 .
Lascano R. J. , Baumhardt R. L. , and Lipe W. N. 1992 Measurement of water flow in young grapevines using the stem heat balance method. Am. J. Enol. Vitie 43 : 159 – 165 .
Lee S. H. , Chung G. C. , Cho B. H. , Guh J. O. , and Suh S. R. 2002 Nutrient deprivation affects xylem sap flow and water channel function in tomato plants. Journal of Plant Nutrition 25 : 1407 – 1413 .
Li Y. , Fuchs M. , Cohen S. , Cohen Y. , and Wallace R. 2002 Water uptake profile response of corn to soil moisture depletion. Plant, Cell and Environment 25 : 491 – 500 .
Lu P. , Woo K. C. , and Liu Z. T. 2002 Estimation of whole-plant transpiration of bananas using sap flow measurements. J. of Exp. Botany 53 : 1771 – 1779 .
Monje O. A. and Bugbee B. 1992 Inherent limitations of nondestructive chlorophyll meters: a comparison of two types of meters. Hortscience 27 : 69 – 71 .
Morgan C. L. S. , Norman J. M. , and Lowery B. 2003 Estimating plant-available water across a field with an inverse yield model. Soil Science Society of American Journal 67 : 620 – 629 .
Nageswara Rao R. C. , Talwar H. S. , and Wright G. C. 2001 Rapid assessment of specific leaf area and leaf nitrogen in peanut (Arachis hypgaea L.) using a chlorophyll meter. J. Agronomy and Crop Science 186 : 175 – 182 .
Nageswara Rao R. C. , Udaykumar M. , Farquhar G. D. , Talwar H. S. , and Prasad T. G. 1995 Variation in carbon isotope discrimination and its relationship to specific leaf area and ribulose-1,5-bisphosphate carboxylase content in groundnut genotypes. Australian Journal of Plant Physiology 22 : 545 – 551 .
Nageswara Rao R. C. and Wright G. C. 1994 Stability of the relationship between specific leaf area and carbon isotope discrimination across environments in peanut. Crop Science 34 : 98 – 103 .
Phene C. J. , Hutmacher R. B. , El Ayars J. , Davis K. R. , Mead R. M. , and Schoneman R. A. 1992 Maximizing water use efficiency with subsurface drip irrigation. ASAE Paper No. 92-2090 St. Joseph, Mich ASAE .
SAS 1997 JMP Statistical Discovery Software Cary, NC, USA SAS Institute Inc .
Senock R. S. and Ham J. M. 1995 Measurements of water use by prairie grasses with heat balance sap flow gauges. J. Range Management 48 : 150 – 158 .
Senock R. S. , Ham J. M. , Loughin T. M. , Kimball B. A. , Hunsaker D. J. , Pinter P. J. , Wall G. W. , Garcia R. L. , and LaMorte R. L. 1996 Sap flow in wheat under free-air CO2 enrichment. Plant, Cell and Environment 19 : 147 – 158 .
Sorensen R. B. , Jones T. L. , Campbell G. S. , and Montes-Helu M. 1999 Heat pulse needles to measure pecan tree transpiration. ASAE 15 : 651 – 657 .
Sorensen R. B. , Wright F. S. , and Butts C. L. 2001a Pod yield and kernel size distribution of peanut produced using subsurface drip irrigation. Applied Engineering in Agriculture 17 : 165 – 169 .
Stansell J. R. , Shepherd J. L. , Pallas J. E. , Bruce R. R. , Minton N. A. , Bell D. K. , and Morgan L. W. 1976 Peanut responses to soil water variables in the southeast. Peanut Science 3 : 44 – 48 .
Tanner C. B. 1963 Plant temperatures. Agron. J 55 : 210 – 211 .