


Introduction
Germplasm collections are an invaluable source for crop improvement. They include diverse landraces, obsolete and modern cultivars, breeding lines, germplasm lines, genetic stocks and crop wild relatives (Dempewolf et al., 2017; Cockel et al., 2022; Tirnaz et al., 2022; Lazaridi et al., 2024). They contain a vast amount of genetic variability for many agronomically desirable traits such as increased yield, improved disease resistance, enhanced drought tolerance, superior nutritional quality and other processing attributes. This genetic variability provides raw materials to breeders for developing improved cultivars (Haussmann et al., 2004; Halewood et al., 2018; Swarup et al., 2021; Ghamkhar et al., 2025). Many of the world’s crop germplasm collections are maintained in public genebanks at regional, national and international centers. It was estimated that there are about 1,750 genebanks worldwide with about 7 million accessions conserved (Khoury et al., 2022).
The United States Department of Agriculture (USDA) peanut germplasm collection was assembled over eight decades and is securely managed at the National Plant Germplasm System (NPGS) genebank in Griffin, GA. The germplasm collection is a testimony to the vision and dedication of several prominent national and international collectors with funding support from the USDA, the International Board for Plant Genetic Resources (IBPGR), UN Food and Agriculture Organization (FAO), Consultative Group on International Agricultural Research (CGIAR), the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) and logistic support from host countries. Substantial support was also provided to the different collectors from their respective institutions/agencies. The germplasm collection contains about 9,000 cultivated peanut (Arachis hypogaea L.) accessions and 600 wild Arachis species accessions. The accessions were obtained from germplasm collection expeditions and as donations from several countries. About half of the A. hypogaea germplasm collection includes locally adapted types called landraces from the centers of origin and diversity in South America (Holbrook, 2001; GRIN Global). These landraces accumulated adaptation genes for survival and propagation under extremely varied climatic conditions with a wide range of biotic and abiotic stresses. The other half of the collection was acquired from countries outside of South America with a third of the collection from several countries in Africa (Holbrook, 2001; GRIN Global). The collection represents a tremendous amount of morphological variability for many plant, pod and seed traits (GRIN Global; Simpson et al., 1992; Wang et al., 2022). The first peanut Plant Introduction (PI) was PI 4253 from Cairo, Egypt (Isleib et al., 2001). It was donated by B. Lathrop and D. G. Fairchild in 1899 and is no longer available. However, the oldest and still an actively distributed A. hypogaea accession is, PI 109839, collected in Venezuela in 1935 (GRIN Global). PI 109839 has leaf spot resistance (Foster et al., 1980) and is still an active accession in the genebank for distribution. Additionally, the wild Arachis species germplasm collection in the USDA genebank represents a unique genepool for many economically useful traits that are not present within the cultivated peanut, thus providing novel genetic variability to expand the genetic base of A. hypogaea (Stalker et al., 2013; Stalker, 2017). All wild species accessions collected in South America were spread across Argentina, Bolivia, Brazil, Paraguay and Uruguay. The first wild Arachis species acquired into the USDA genebank is A. glabrata in 1936 (GRIN Global).
Although the USDA peanut germplasm collection is large with approximately 9,600 accessions, the acquisition of new germplasm, especially from the centers of origin and diversity in South America, would greatly enhance the value of the collection for future research. Native peanut genetic resources in several countries of South America are being eroded due to deforestation, cattle ranching and urbanization activities (Baumann et al., 2017; de La Sancha et al., 2021; Mosciaro et al., 2023) and must be rescued for ex situ conservation before they become extinct. However, international laws governing germplasm access and use have limited the efforts to acquire additional peanut germplasm from South America. Considering this and other anthropological threats, the maintenance and availability of the USDA germplasm collection is extremely critical for the long-term sustainability of the U.S. peanut industry.
The objectives of this paper are to describe the activities associated with management of the USDA peanut germplasm collection, highlight its importance to the U.S. peanut industry, discuss the different international treaties and their implications for germplasm access, use, and benefit sharing, and provide recommendations for future germplasm collection, conservation and access.
Status, maintenance and management of the USDA peanut germplasm collection
Status of the germplasm collection
The peanut germplasm collection is a national as well as a global treasure. Researchers within the U.S. and from across the globe regularly request germplasm from the collection. As of December 2025, the collection has 8,940 and 622 accessions of A. hypogaea and wild Arachis species, respectively. The A. hypogaea collection includes accessions from 97 countries and represents both subspecies, hypogaea and fastigiata, and the six botanical varieties within the subspecies (GRIN Global). The wild Arachis species collection includes accessions from 69 of the 84 species from South America (GRIN Global). The availability of the germplasm collection for research and educational uses is an important objective of the genebank. About 8,222 A. hypogaea accessions and 417 accessions of the wild Arachis species are currently available for distribution. Many wild species produce limited quantities of pods during regenerations. As such, efforts are underway to increase their availability for research collaborators. In 2025, 2,208 accessions of A. hypogea were distributed for research use. Of these, 2,035 were sent to domestic users and 173 to international researchers. Similarly, 156 accessions of wild Arachis species were distributed with 144 for domestic research use and 12 to international researchers. Another important goal is to securely back up the germplasm collection at a secondary site. About 99% of the A. hypogaea collection and 74% of the wild Arachis species accessions were backed up at the National Laboratory for Genetic Resources Preservation (NLGRP) in Fort Collins, CO.
Maintenance of the germplasm collection
The A. hypogaea germplasm collection is maintained for long-term storage at -18oC. Up to 400 seeds of each accession (PI) are stored in tightly packed, heat-sealed aluminum foil bags. A sample of each PI is also maintained at 4oC temperature and 25% relative humidity for short-to-medium term storage in regular coffee paper bags. This serves as the distribution sample. When researchers request seeds of a specific germplasm line, seeds from the 4oC sample are taken to package and ship. If the on-hand seed quantity of a PI falls below 200, they become unavailable for distribution. Similar procedures are followed to maintain wild Arachis species accessions except that each accession is maintained as pods.
Management of the germplasm collection
Although the accessions are securely stored and maintained, they require periodic regenerations to replenish fresh seeds into the genebank. Regular seed viability tests for peanut are conducted following the protocols of the Association of Official Seed Analysts (AOSA, 1981). The viability of freshly regenerated seeds before they are processed into the collection is determined for a base line seed viability. Then, randomly selected accessions maintained at -18oC are tested after 5, 10 and 15 years of storage to monitor viability. If the seed viability drops to 50% or less, the accessions are prioritized for regeneration. Most seeds remain viable for 15 or more years in -18oC storage. Annually, between 600 and 800 A. hypogaea accessions are regenerated in field plots. Close attention is paid during seed packaging, and care is taken during planting and harvesting to avoid inadvertent mix-ups to maintain the purity and integrity of the PIs. Digital images of seeds are captured before planting and compared to the harvested materials to make sure they match, as an additional check to maintain the purity of the PIs. The germplasm collection displays significant morphological genetic variability for many plant, pod and seed features (GRIN Global). During the regeneration process, a large amount of descriptor data is collected to characterize the germplasm for several phenotypic traits (Pittman, 1995) including digital images of pods and seeds. The A. hypogaea collection is also characterized for important agronomic traits including nutritional components to identify desirable germplasm for use in peanut breeding programs (GRIN Global; Wang et al., 2022). The descriptor data is available to researchers at the GRIN Global public website, https://npgsweb.ars-grin.gov/gringlobal/search.
Additionally, about 100 wild Arachis species accessions are regenerated annually. Wild species are grown in containers in the greenhouses for ease of maintenance and harvest. Wild species are relatively difficult to grow, and not all species produce enough pods in one growing season, with some taking several years of regenerations to obtain even 100 pods. The containers are checked regularly to make sure that the branches of the plants are confined to the same container and do not grow out into another nearby container of a different species. Leal-Bertioli et al. (2024) characterized section Arachis species accessions from the USDA genebank collection along with the same accessions from other wild Arachis species germplasm collections for molecular genetic variability and developed molecular diagnostic tools for the positive identification of section Arachis species accessions. This is a valuable resource to characterize accessions of unknown taxonomic classification in the USDA germplasm collection to assign to the corresponding species within section Arachis.
Brief history of the USDA peanut germplasm collection
Systematic efforts to assemble the peanut germplasm collection started in the 1930’s and below is a historical summary of the collection as retrieved from GRIN Global. Additional historical information on Arachis germplasm collections can be found in Gregory et al. (1973); Banks (1976); Simpson (1984); and Krapovickas and Gregory (1994 and 2007).
The USDA peanut germplasm collection contained 50 A. hypogaea accessions and one wild Arachis species (A. glabrata) accession in 1936 (GRIN Global). Twenty years later (1950-59), the germplasm collection grew to 1,195 A. hypogaea accessions and 18 wild species accessions. This was largely possible due to the efforts of the USDA supported germplasm collection trips in collaboration with scientists from Argentina and Brazil as described by Gregory et al. (1973), Simpson (1984), and Krapovickas and Gregory ( 1994; 2007). By the end of the next decade (1960-69), the germplasm collection increased to 3,838 A. hypogaea accessions and 92 wild species accessions (Banks, 1976). Then, between 1976 and 1983, 17 germplasm collection trips were undertaken (Simpson, 1984; Krapovickas and Gregory, 1994; 2007) and by the end of 1989, the peanut germplasm collection contained 7,697 and 255 of A. hypogaea and wild species accessions, respectively. The germplasm collection increased to 8,626 A. hypogaea accessions and 482 wild species accessions before additional acquisitions of international materials, especially from South America, dropped significantly due to the introduction of international laws regulating germplasm access and use in 1993 as discussed in the International treaties for germplasm access, use and benefit sharing section below. Presently, the collection contains about 9,000 A. hypogaea accessions and 600 wild species accessions. The increases of A. hypogaea germplasm are mainly from domestic materials and the additional wild species accessions were donated by NC State University and Texas A&M AgriLife Research.
Importance of the germplasm collection to the U. S. peanut industry
The germplasm collection possibly contains sufficient genetic diversity for many agronomic traits for sustained genetic improvement of peanut. Growers have been demanding resilient cultivars to grow peanuts profitably, especially those without irrigation capabilities to grow the crop. Additionally, persistent disease pressures are increasing costs of fungicide sprays to control them. Peanut breeders have been characterizing the collection for useful traits such as drought tolerance and disease resistances for resilient cultivar development. Below sections highlight the importance and contribution of the germplasm collection to the U.S. peanut industry.
Germplasm contribution to U. S. peanut cultivars
Large genetic variation for economically important traits was observed in the peanut germplasm collection (Holbrook and Isleib, 2001; Isleib et al., 2001; Wang et al., 2022) and it has contributed to the development of several important cultivars in the U. S. It was estimated that the PIs contributed about 20.6% of the ancestry of the cultivars (Isleib and Barkley, 2013). The PI 203396, collected in Porte Alegre, Brazil in 1952, has contributed significantly to Tomato spotted wilt virus (TSWV) and leaf spot resistance in the U. S. cultivars. Thirty-three cultivars released between 1984 and 2017 contained PI 203396 as an ancestor (Branch, 1996; Branch, 2007; Holbrook et al., 2008; Branch and Brenneman, 2015; Tallury et al., 2023; GRIN Global). The economic impact of TSWV resistance from PI 203396 was estimated at $200 million annually (Holbrook, 2001; Isleib et al., 2001). Additionally, selected germplasm was evaluated for resistances to leaf spots, sclerotinia blight, stem rot and peanut smut to develop multiple disease resistant cultivars (Damicone et al., 2010; Branch and Brenneman, 2020; Holbrook et al., 2008, 2017, , 2021, 2023; Bennett et al., 2018; Bennett and Chamberlin, 2020; Massa et al., 2021; Cason et al., 2022; Chamberlin et al., 2022).
The wild Arachis species have been utilized for peanut improvement, especially for developing disease resistant germplasm and cultivars (Stalker and Simpson, 1995; Stalker et al., 2013; Stalker, 2017). The only source of Root Knot Nematode (RKN; Meloidogyne arenaria (Neal) Chitwood race 1) resistance in the currently grown U.S. cultivars is from the wild Arachis species, A. cardenasii Krapov. & W. C. Greg. (Simpson and Starr, 2001; Simpson et al., 2003; Baring et al., 2023; Holbrook et al., 2008, 2017, and 2023). Additionally, several germplasm lines and cultivars derived from wild species with resistance to leaf spots, RKN, and TSWV have been released (Stalker, 1993; Simpson et al., 1993; Stalker et al., 2002a and 2002b; Isleib et al., 2011; Branch and Brenneman, 2015; Bertioli et al., 2021a and 2021b; Cason et al., 2022; Chu et al., 2022; Godoy et al., 2022; Holbrook et al., 2008, 2017, , 2021, 2023; Tallury et al., 2014; Stalker, 2017; Newman et al., 2023; Stalker et al., 2025). Also, neo-allotetraploids were produced to develop new interspecific hybrid derived populations for additional disease evaluations and agronomic performance (Bertioli et al., 2021b; Leal-Bertioli et al., 2021; Leal-Bertioli et al., 2025).
Genetic variability of seed quality traits for nutritional improvement
Wang et al. (2022) analyzed about 8,400 A. hypogaea PIs from the USDA genebank collection for several seed quality traits of importance to the peanut industry. The traits included 100-seed weight, seed coat color, total oil, and fatty acid profiles. They reported that the 100-seed weight ranged from 22 g in PI 270904 to 160 g in PI 221068 with an overall mean weight of 49 g. Among the PIs, a large amount of variation in seed coat colors was observed with 2,552 Tan, 2,442 Pink, 2,221 Red, 678 Purple, 148 White and 425 Multicolor types, respectively. Recently, the peanut industry expressed an interest in high oil types for the export market. Again, Wang et al. (2022) demonstrated significant variation for total oil among the 8,400 PIs with oil content ranging from 37% in PI 487337 (var. vulgaris) to 59% in PI 668529 (var. hypogaea) with a mean oil content of 49%. They also indicated variation in oleic and linoleic acids with means of 45% and 33%, respectively. Oleic acid ranged from 31% to 82% whereas linoleic acid ranged from 2% to 47% among the samples analyzed. Also, Wang et al. (2013) evaluated the U.S. peanut mini-core accessions and reported significant variability for seed chemical composition including for oil, fatty acid profiles, quercetin and seed weight. Later, Wang et al. (2015) identified two new high-oleic acid mutants, PI 342664 and PI 342666, belonging to the A. hypogaea subsp. hypogaea which provide unique sources of new germplasm for peanut breeding research. Many of these PIs are potential sources for peanut nutritional improvement and development of healthy snacks or food items.
Wild Arachis species have been used for food, forage and as ornamental ground cover (Stalker 2017). Wang et al. (2010) reported significant variation for oil content among 39 wild Arachis species in the USDA germplasm collection with A. magna containing 61% compared to 56% in the A. hypogaea check. Similarly, Upadhyaya et al. (2011) identified several wild Arachis species accessions with useful agronomic and nutritional traits, including high sugar, high protein and high oil content. Further, Grosso et al. (2000) conducted a detailed seed nutritional characterization study including total oil, fatty acid composition, ash, protein, carbohydrate and sterol contents of 17 wild Arachis species. Though they reported a large variation among the species for the chemical components, it was concluded that none were superior to the cultivated peanut for enhancing its nutritional composition. The USDA perennial peanut germplasm collection of A. glabrata was used to release 10 peanut cultivars primarily for forage use (Shahid et al., 2023). The cultivars are also promoted as a groundcover in orchard farms for ease of maintenance, aesthetics, biological nitrogen fixation ability and as an ecosystem for beneficial insect pollinators (Shahid et al., 2023).
Emerging and evolving threat to the U. S. peanut industry
Peanut smut, caused by the soil borne fungus, Thecaphora frezzii Carranza & Lindquist, is an emerging threat to the U. S. peanut industry. Currently, it is present in 100% of the commercial seed production farms in Argentina causing crop losses of up to 35% and also found in Bolivia and Brazil on wild Arachis species (Rago et al., 2017). However, with increased global trade and travel, it is possible for the pathogen to spread beyond these borders. Should this occur, the U.S. peanut industry would be at risk of collapse without resistant cultivars. Fungicides used for controlling common peanut diseases have little or no effect on the smut fungus. Consequently, developing resistant cultivars is the most logical option to protect the crop from smut (Arias et al., 2019; de Blas et al., 2019; Kearney et al., 2021). USDA researchers at the National Peanut Research Laboratory, Dawson, GA; Wheat, Peanut and Other Field Crops Research Unit, Stillwater, OK; Crop Genetics and Breeding Research Unit, Tifton, GA; and the Plant Genetic Resources Conservation Unit, Griffin, GA, in collaboration with the U.S. peanut industry and Argentinian scientists, have been proactively identifying smut resistant germplasm for cultivar development. They are also investigating the ecology and genetics of the pathogen to understand the different pathotypes, genetic variability and patterns of spread of the smut fungus in South America to develop appropriate management practices for controlling the disease (Arias et al., 2019; Arias et al., 2023). Because of access to diverse PIs in the genebank, the U. S. researchers evaluated a select set of germplasm including some popular cultivars and other breeding materials in heavily smut infested fields over multiple years in Argentina. This led to the identification of eight PIs as sources of peanut smut resistance (Chamberlin et al., 2022). These PIs were used in crosses with U.S. cultivars to develop populations for genetic mapping studies to develop smut resistant cultivars quickly and efficiently using molecular assisted breeding strategies (Chamberlin et al., 2024). In a related study, Massa et al. (2021) analyzed five additional PIs from the USDA peanut germplasm collection which are closely related to a smut resistant Bolivian landrace to verify if the PIs contained resistance. Their study not only confirmed resistance but also traced the resistance in the PIs to the landrace. Additionally, de Blas et al. (2019) demonstrated smut resistance in wild Arachis species and produced a resistant amphidiploid. It was crossed with A. hypogaea to develop a recombinant inbred line (RIL) population with 22 highly resistant RILs, thus illustrating the incorporation of smut resistance from the wild Arachis species into A. hypogaea (de Blas et al., 2019). These studies demonstrate that the genetic variability conserved in the peanut germplasm collection provides insurance against unforeseen threats such as smut to the U.S. peanut production. It also highlights the importance and value of germplasm conservation, maintenance, and utilization for peanut improvement.
International treaties for germplasm access, use and benefit sharing:
Historically, germplasm collected anywhere in the world was freely shared as a “Global Public Goods” with free access for crop improvement. However, in the 1980s, concerns about the loss of biodiversity became widespread and fueled fears that germplasm resources may be privatized and the derived commercial products sold back to the countries of origin of the germplasm. These fears led to a movement of advocating for national sovereignty of biodiversity and for mechanisms where a portion of the monetary benefits derived from the use of biodiversity would flow back to the country of origin of the germplasm. As a result, a new biodiversity framework was established (Williams and Williams, 2001). The United Nations Environment Program (UNEP) developed the Convention on Biological Diversity (CBD) (United Nations, 1992) and its associated “Nagoya Protocol” (Secretariat of the Convention on Biological Diversity, 2011). These new governance frameworks, that covered all biological diversity of microbial, animal and plants, intended to promote biodiversity conservation, access and equitable sharing of benefits. Later, because of the importance of plant genetic resources (PGRs) for crop improvement and global food security, the International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA) or Plant Treaty (PT), administered by the UN Food and Agriculture Organization (FAO), came into force on 29th June 2004 (FAO, 2002). These two treaties, together with the Nagoya Protocol, provided the basic framework with responsibilities and obligations for access, use and benefit sharing of the global germplasm resources (Marden et al., 2023). Below is a brief description of each treaty.
Convention on Biological Diversity (CBD)
The CBD (https://www.cbd.int) came into force on 29th December 1993, aimed to promote conservation, sustainable use of biological diversity (microbial, animal and plant) and encourage equitable sharing of the benefits resulting from the use of the genetic resources via the Nagoya Protocol. Benefit sharing was viewed as an incentive for continued and sustained conservation of biological diversity. Additionally, the CBD recognized that countries held sovereignty over their native genetic resources and their right to provide access to it. An extreme interpretation of these claims led several countries, including those containing Arachis genetic resources, to implement national laws regulating their access and use. This has hindered access to peanut genetic resources from the centers of origin and diversity in south and central America (Williams and Williams, 2001). The CBD is a bilateral treaty, with negotiation of germplasm access agreed between the two countries, the provider country containing the native germplasm and the country that is receiving the germplasm. The implementation of CBD has led to laborious and burdensome regulations to access germplasm resources with prior information consent (PIC) and mutually agreed terms (MAT) for sharing of benefits. Also, countries established their own interpretations and additional regulations, thus imposing further restrictions on germplasm access (Brink and van Hintum, 2020; Williams et al., 2020). Ironically, these laws have restricted germplasm access to researchers in south and central America as well. The CBD was ratified by 196 countries, and the U.S.A. is not one of them.
Plant Treaty (PT) or International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA)
The objectives of the PT (https://www.fao.org/plant-treaty/en/) are similar to those of the CBD of conservation and sustainable use of genetic resources, especially of plant genetic resources (PGRs) important for food and agriculture (PGRFA). The PT offered a compromise framework to member states to contribute any or all of the PGRs originated in their countries into a pool to share with the rest of the member countries. Also, a portion of any financial benefits resulting from their use would be deposited into a fund to help with continued conservation and other sustainable agricultural production practices for global food security. It also recognized that no country is self-sufficient in PGRs of major food crops and that countries are interdependent (Khoury et al., 2022). Consequently, it advocated collective conservation for long-term sustainability of PGRs under a multilateral system (MLS). MLS is a global pool of PGRFA of 64 important food and forage crops which are included in the Annex 1 list of crops of the treaty (FAO, 2002). These crops account for 80% of humanity’s food supply. Paradoxically, peanut was excluded from the Annex 1 list, which practically shut the door for new germplasm access from South America, the home of peanut origin and diversity. As of 1st April 2025, the PT has 154 members including the U.S.A.
Although far from perfect, the PT has simplified international access of PGRs to genebanks and other public-sector researchers. It provides access to germplasm resources through a Standard Material Transfer Agreement (SMTA). Most of the USDA National Plant Germplasm System (NPGS) accessions are included in the MLS.
World Intellectual Property Organization (WIPO) Treaty
In May 2024, the World Intellectual Property Organization (WIPO) Treaty on Intellectual Property, Genetic Resources and Associated Traditional Knowledge was adopted by the member states. The treaty aims to enhance transparency of the patent system via a database of genetic resources and associated traditional knowledge of indigenous peoples and local communities. The database will be accessible to patent offices to prevent patents being granted erroneously for inventions that are not novel with regard to genetic resources and traditional knowledge or information. Evidently, this is the first WIPO Treaty to address the interface between intellectual property, genetic resources and traditional knowledge and included benefit sharing, specifically for indigenous peoples as well as local communities. Again, the benefit sharing is deemed as an incentive for indigenous peoples and communities to safeguard the native germplasm resources.
Although the treaties described above have the common goal of germplasm conservation, access for sustainable use and sharing of benefits, they also impose burdensome regulations for access and use, thus defeating the very purpose of the treaties.
Implications of the treaties for the USDA peanut germplasm collection
Since peanut was not among the Annex 1 list of crops of the multilateral system of PT, it has become impossible to acquire new germplasm from South America for entry into the USDA genebank collection. Charles Simpson, Texas A&M AgriLife Research, Stephenville, TX, has been helping with restoration and replenishment of the peanut germplasm collection in the USDA genebank since the 1980s and is still assisting the peanut curator with management of the germplasm collection. He participated in additional germplasm collection trips in Brazil until 2016 but none of those materials, new or old, were allowed to be taken out of Brazil because of the international treaty restrictions. Moreover, there are several indigenous A. hypogaea types in south and central America which are grown and consumed by local communities that could be valuable additions to the germplasm collection and breeding programs. Unfortunately, some of these types are being eroded because of urbanization, cattle ranching activities and other anthropogenic actions (Baumann et al., 2017; de La Sancha et al., 2021; Mosciaro et al., 2023). Further, there are geographical and taxonomic gaps in the genebank collection and acquisition of materials to fill these gaps would greatly enhance the USDA peanut germplasm collection for future research (GRIN Global). Halewood et al. (2012) mentioned that global germplasm exchanges decreased after the implementation of the CBD due to uncertainty of legal implications, restrictive national policies, and/or countries unwilling to share their genetic resources without being included in royalties or some form of renumeration. The USDA peanut germplasm collection experienced a similar trend with overall reductions in germplasm acquisition post-CBD.
Khoury et al. (2022) articulated that if a crop becomes a major producer outside of its primary origin and region(s) of diversity, then there is significant interdependence globally for genetic resources from the centers of origin and diversity for its genetic improvement. Peanut is predominantly cultivated in many countries outside of the crop’s primary centers of origin and diversity in South America. As a result, peanut-producing countries are dependent on genetic resources for crop improvement. Because of the international treaty restrictions, peanut germplasm resources from South America became inaccessible to many countries. Hence, it is even more important to continue to maintain the USDA peanut germplasm collection in the best possible condition for sustained domestic production.
Recommendations for future germplasm conservation and collection
The recommendations below must be coordinated by the secretariats of CBD, PT, Crop Trust and other international agencies for possible funding support for the different activities. Additionally, recent success stories of international collaborations for the benefit of global peanut research community must be highlighted. The U. S. peanut industry coordinated these efforts and provided a major part of the financial support. One of the success stories is that of the International Peanut Genome Initiative (IPGI). IPGI demonstrated that joint international research and collaborations are possible for the benefit of the global peanut research community with the development of peanut genome sequences. The other includes the collaboration between the U. S. and Argentinian researchers that peanut smut resistant germplasm and associated information will be available globally if the disease ever expands beyond the borders of South America.
Address the reasons for non-inclusion of peanut in the Annex 1 list of crops
In his detailed assessment of the global strategy for the conservation and use of peanut genetic resources, Williams (2022), observed that the absence of peanut and its wild relatives in the Annex 1 list of crops of the PT was a serious bottleneck for international exchange of peanut genetic resources and advocated that continued dialogue with the respective member countries must be pursued to resolve this gridlock. Further, in their classic paper on the legacy genetics of A. cardenasii, Bertioli et al. (2021a) provided a strong impetus for free and unrestricted access to peanut germplasm and its benefits for global peanut production. They elegantly demonstrated the benefits of one Arachis species accession to develop leaf spot resistant germplasm for sustainable production globally. They also emphasized the negative consequences imposed by the existing international treaties to germplasm access and highlighted the need and value of free exchange of germplasm for global peanut improvement. Such examples may help members of CBD and PT to reconsider the inclusion of peanut in Annex 1 list of crops.
International initiatives to protect peanut genetic variability in situ and ex situ
As described elsewhere in this chapter, anthropogenic activities are leading to the rapid loss and extinction of native genetic variability in south and central America in the primary centers of origin and diversity as well as in the secondary centers of diversity in Africa and Asia (Baumann et al., 2017; de La Sancha et al., 2021; Chapman et al., 2022; Mosciaro et al., 2023; Yang et al., 2023). As shown in Fig. 1, the effects of human population growth and urbanization have led to the extinction of the A. duranensis population, the female genome donor of A. hypogaea, in Argentina. Fortunately, the A. duranensis accession from this population is still maintained in several genebanks including the USDA genebank. While wild peanut populations in native habitats are experiencing genetic erosion due to limited or lack of effective in situ conservation strategies, the ex situ conserved germplasm is being lost due to insufficient physical and technical resources for its maintenance. Many peanut researchers in South America understand the gravity of this issue as well. Reversing these activities must be an urgent and immediate goal of the global peanut research community before more valuable germplasm is lost. A logical recommendation is that regional initiatives must be promoted among the neighboring countries to highlight the common benefits of PGRs, their conservation, management and access to use (Tyagi, 2022).
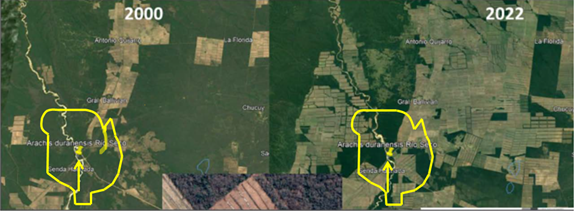
Site of A. duranensis, the female genome donor of A. hypogaea, in Río Seco, Salta, Argentina in 2000 and 2022 indicatd by the yellow arrows. In 2022, the surrounding areas indicated by yellow boundaries were taken over by developments and leading to the loss of the original population.
Explore with south and central American countries to reopen germplasm collection and exchange
Williams (2022) stated “if the current treaty restrictions had existed in the past, there wouldn’t be any global collections of peanut germplasm conserved in genebanks around the world, as there are now”. Past USDA Arachis germplasm explorations in South America provided non-monetary benefits to the host country with supporting the travel expenses of their personnel on the trip, any equipment costs of the exploration, sharing half of the collected germplasm, preparation of herbarium specimens, and authorship in publications of research (Williams and Williams, 2001). Such incentives helped all parties involved in the collection trips to understand and develop personal relationships with each other and led to many productive collaborations. Similar incentives must be pursued to revive germplasm collection and train the next generation of germplasm curators and managers. Undeniably, capacity building and training would greatly benefit with rescuing threatened germplasm resources.
Highlight global benefit of conserving germplasm resources at multiple regions
The threats of unexpected biological events such as new pathogens or insect pests, drought, and mechanical breakdowns could destroy the in situ as well as ex situ collections. Following the implementation of the CBD, germplasm exchanges became increasingly restricted, even among the south and central American countries. Thus, it is critical to preserve the existing ex situ collections, and any new germplasm collected at more than one location/region. The USDA genebank is equipped with technical and physical resources to safeguard the peanut germplasm collection with secured back up at the National Laboratory for Genetic Resources Preservation (NLGRP) in Fort Collins, CO.
Summary
The USDA peanut germplasm collection offers the essential genetic resources for researchers to develop improved cultivars. The germplasm collection possibly contains sufficient genetic diversity for sustained peanut improvement and is considered a national as well as a global treasure. It has contributed to several important cultivars and continues to. The collection was assembled over many decades of germplasm expeditions and donations from other countries. However, with the Convention on Biological Diversity and the exclusion of peanut from the Plant Treaty Annex 1 list of crops, acquisition of new germplasm from South America stalled. Added to this, anthropogenic activities are leading to the extinction of native peanut genetic resources in the crop’s centers of origin and diversity and secondary centers of diversity elsewhere. There are several success stories from free and unrestricted germplasm exchanges as highlighted in this paper. The global peanut research community must act now to reverse the trend for sustained in situ and ex situ conservation and use of these valuable genetic resources for sustained peanut improvement.
Disclaimer
The authors declare no conflict of interest. Material presented in this paper is solely for the purpose of providing specific scientific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Literature Cited
AOSA. 1981. Rules for testing seeds. Arachis hypogaea. J. Seed Tech. 6:36-38.
Arias R.S., Cazon L.I., Massa A.N., Scheffler B.E., Sobolev V.S., Lamb M.C., Duke M.V., Simpson S. A., Conforto C., Paredes J. A. and Buteler M. I.. 2019. Mitogenome and nuclear-encoded fungicide-target genes of Thecaphora frezii-causal agent of Peanut smut. Fungal Genom. Biol. 160:1–8.
Arias R.S., Conforto C., Orner V.A., Carloni E.J., Soave J.H., Massa A.N., Lamb M.C., Bernardi-Lima N. and Rago A.M.. 2023. First draft genome of Thecaphora frezii, causal agent of peanut smut disease. BMC Genomic Data. 24:9. doi: [: 10.1186/s12863-023-01113-w].
Banks D.J. 1976. Peanuts: Germplasm resources. Crop Sci. 16:499-502.
Baring M.R., Cason J.M., Burow M.D., Simpson C.E., Chagoya J. and Bennett B.D.. 2023. Registration of ‘NemaTAM II’ peanut. J. Plant Reg. 17:291-298.
Baumann M., Israel C., Piquer-Rodríguez M., Gavier-Pizarro G., Volante J.N. and Kuemmerle T.. 2017. Deforestation and cattle expansion in the Paraguayan chaco 1987–2012. Regional Environmental Change. 17:1179-1191.
Bertioli D.J., Clevenger J., Godoy I.J., Stalker H.T., Wood S., Santos J.F., Ballen-Taborda C., Abernathy B., Azevedo V., Campbell J., Chavarro C., Chu Y., Farmer A.D., Fonceka D., Gao D., Grimwood J., Halpin N., Korani W., Michelotto M.D., Ozias-Akins P., Vaughn J., Youngblood R., Moretzsohn M.C., Wright G.C., Jackson S.A., Cannon S.B., Scheffler B.E. and Leal-Bertioli S.C.M.. 2021a. Legacy genetics of Arachis cardenasii in the peanut crop shows the profound benefits of international seed exchange. Proc. Natl. Acad. Sci. U S A. 118(38):e2104899118. doi: [: 10.1073/pnas.2104899118].
Bertioli D. J., Gao D., Ballen-Taborda C., Chu Y., Ozias-Akins P., Jackson S.A., Holbrook C.C. and Leal-Bertioli S.C.M.. 2021b. Registration of GA-BatSten1 and GA-MagSten1, two induced allotetraploids derived from peanut wild relatives with superior resistance to leaf spots, rust, and root-knot nematode. J. Plant Reg. 15:372-378.
Bennett R., and Chamberlin K.. 2020. Resistance to Athelia rolfsii and web blotch in the U.S. mini-core collection. Peanut Sci. 47:17-24. doi: [: 10.3146/PS19-18.1].
Bennett R.S., Chamberlin K. D. and Damicone J.P.. 2018. Sclerotinia blight resistance in the US peanut mini-core collection. Crop Sci. 58:1306-1317.
Branch W.D. 1996. Registration of ‘Georgia Green’ peanut. Crop Sci. 36:806.
Branch W.D. 2007. Registration of ‘Georgia-06G’ peanut. J. Plant Reg. 1:120.
Branch W.D. and Brenneman T.B.. 2015. Registration of ‘Georgia-14N’ peanut. J. Plant Reg. 9:159-161.
Branch W.D. and Brenneman T.B.. 2020. Registration of ‘Georgia- 19HP’ peanut. J. Plant Reg. 14:306-310.
Brink M., and van Hintum T.. 2020. Genebank operation in the arena of access and benefit-sharing policies. Frontiers in Plant Sci. 10:1712.
Cason J.M., Simpson C.E., Burow M.D., Tallury S., Pham H. and Ravelombola S.W.. 2022. Use of wild and exotic germplasm for resistance in peanut (Arachis hypogaea L.). J. Plant Reg. 17:1-25.
Chamberlin K.D., Baldessarri J., Bennett R.S., Clevenger J. P., Holbrook C. C., Tallury S. P., Chu Y., Ozias-Akins P., Conde M. B. and Payton M. E.. 2022. Identification of germplasm resistant to peanut smut. Peanut Sci. 49:1-16. doi: [: 10.3146/0095-3679-491-PS21-10].
Chamberlin K., Bennett R., Baldessarri J., Barrera G., Cordes G., Grandon N., Mamani E., Rodriguez A., Morchetti S., Holbrook C., Ozias-Akins P., Chu Y., Tallury S., Clevenger J., Korani W., Scheffler B., Youngblood R. and Simpson S.. 2024. Discovery of a resistance gene cluster associated with smut resistance in peanut. Peanut Sci. 51:59-65. doi: [: 10.3146/0095-3679-51-PS23-6].
Chapman C.A., Abernathy K., Chapman L.J., Downs C., Effiom E.O., Gogarten J.F., Golooba M., Kalbitzer U., Lawes M.J., Mekonnen A., Omeja P., Razafindratsima O., Sheil D., Tabor G.M., Tumwesigye C. and Sarkar D.. 2022. The future of sub-Saharan Africa’s biodiversity in the face of climate and societal change. Front. Ecol. Evol. 10:790552. doi: [: 10.3389/fevo.2022.790552].
Chu Y., Clevenger J.P., Holbrook C.C., Isleib T.G. and Ozias-Akins P.. 2022. Registration of two peanut recombinant inbred lines (TifGP-5 and TifGP-6) resistant to late leaf spot disease. J. Plant Reg. 16:635-640.
Cockel C.P., Guzzon F., Gianella M., and Műller J.V.. 2022. The importance of conserving crop wild relatives in preparing agriculture for climate change. CABI Reviews. doi: [: 10.1079/cabireviews202217031].
Damicone J.P., Holbrook C.C., Smith D.L., Melouk H. A. and Chamberlin K. D.. 2010. Reaction of the core collection of peanut germplasm to Sclerotinia blight and pepper spot. Peanut Sci. 37:1–11. doi: [: 10.3146/PS09-001.1].
de Blas F., Bressano M., Teich I., Balzarini M.G., Arias R.S., Manifesto M.M., Costero B.P., Oddino C., Soave S.J., Buteler M.I., Massa A.N., and Seijo J.G.. 2019. Identification of smut resistance in wild Arachis species and its introgression into peanut elite lines. Crop Sci. 59:1657–1665.
de La Sancha N. U., Boyle S. A., McIntyre N. E., Brooks D.M., Yanosky A., Soto E.C., Mereles F., Camino M., and Stevens R.D.. 2021. The disappearing dry chaco, one of the last dry forest systems on earth. Landscape Ecol. 36:2997-3012.
Dempewolf H., Baute G., Anderson J., Kilian B., Smith C. and Guarino L.. 2017. Past and future use of wild relatives in crop breeding. Crop Sci. 57:1070-1082.
FAO. 2002. International Treaty on Plant Genetic Resources for Food and Agriculture. Rome: Food and Agriculture Organization of the United Nations.
Foster D. J., Wynne J.C. and Beute M.K.. 1980. Evaluation of detached leaf culture for screening peanuts for leafspot resistance. Peanut Sci. 7:98-100. doi: [: 10.3146/i0095-3679-7-2-10].
Ghamkhar K., Hay F.R., Engbers M., Dempewolf H. and Schurr U.. 2025. Realizing the potential of plant genetic resources: the use of phenomics for genebanks. Plant People Planet. 7:23-32.
Godoy I.J., Santos J.F., Moretzsohn M.C., Moraes A.R.M., Michelotto M.D., Bolonhezi D., Nakayama F., de Freitas R.S., Bertioli D.J. and Leal-Bertioli S.C.M.. 2022. ‘IAC SEMPRE VERDE’: a wild-derived peanut cultivar highly resistant to foliar diseases. Crop Breeding and Appl. Biotechnology. 22:e41252232.
Gregory W.C., Gregory M.P., Krapovickas A., Smith B.W. and Yarbrough J.A.. 1973. Structure and genetic resources of peanuts. Chapter 3. pp. 47-134 In Peanuts: Culture and uses. APRES, Stillwater, OK.
GRIN Global. Germplasm Resources Information Network-Global. USDA-ARS. https://npgsweb.ars-grin.gov/gringlobal/search.
Grosso N.R., Nepote V., and Guzmán C.A.. 2000. Chemical composition of some wild peanut species (Arachis L.) seeds. J. Agric. Food Chem. 48:806-809.
Halewood M., Chirugwi T., Hamilton R.S., Kurtz B., Marden E., Welch E., Michiels F., Mozafari J., Sabran M., Patron N., Kersey P., Bastow R., Dorius S., Dias S., McCouch S. and Powell W.. 2018. Plant genetic resources for food and agriculture: opportunities and challenges emerging from the science and information technology revolution. New Phytologist. 217:1407-1419.
Halewood M., Noriega I. L., and Louafi S.. 2012. Crop genetic resources as a global commons: challenges in international law and governance. Abingdon, RoutledgeISBN 9781844078936.
Haussmann B.I.G., Parzies H.K., Presterl T., Sušić Z. and Miedaner T.. 2004. Plant genetic resources in crop improvement. Plant Genet. Res. 2:3-21.
Holbrook C. 2001. Status of the Arachis germplasm collection in the United States. Peanut Sci. 28:84-89. doi: [: 10.3146/i0095-3679-28-2-9].
Holbrook C. and Isleib T.. 2001. Geographical distribution of genetic variability in Arachis hypogaea. Peanut Sci. 28:80-84. doi: [: 10.3146/i0095-3679-28-2-8].
Holbrook C.C., Ozias-Akins P., Chu Y., Brenneman T. B. and Culbreath A.K.. 2023. Registration of ‘TifNV-HG’ peanut. J. Plant Reg. 17:512-516.
Holbrook C.C., Ozias-Akins P., Chu Y., Culbreath A.K., Kvien C.K. and Brenneman T.B.. 2017. Registration of ‘TifNV-High O/L’ peanut. J. Plant. Reg. 11:228-230.
Holbrook C.C., Ozias-Akins P., Chu Y., Lamon S., Bertioli D.J., Leal-Bertioli S.C.M., Culbreath A.K. and Godoy I.. 2021. Registration of TifGP-3 and TifGP-4 peanut germplasm lines. J. Plant Reg. 16:120-123.
Holbrook C.C., Timper P., Culbreath A.K. and Kvien C.K.. 2008. Registration of ‘Tifguard’ peanut. J. Plant Reg. 2:92–94.
Isleib T. and Barkley N.L.. 2013. Contributions of plant introductions to the ancestry of current U.S. peanut cultivars. Abstract. Proc. of Amer. Peanut Res. and Educ. Soc. 58.
Isleib T.G., Holbrook C.C. and Gorbet D.W.. 2001. Use of plant introductions in peanut cultivar development. Peanut Sci. 28:96-113. doi: [: 10.3146/i0095-3679-28-2-11].
Isleib T.G., Milla-Lewis S.R., Pattee H.E., Copeland S.C., Zuleta M.C., Shew B.B., Hollowell J.C., Sanders T.H., Dean L.O., Hendrix K.W., Balota M. and Chapin J.W.. 2011. Registration of ‘Bailey’ Peanut. J. Plant Reg. 5:27-39.
Kearney M., Zuza M., Ibanez M., Peralta V., Peiretti G., Alcalde M., Mojica C. and Rago A.. 2021. Response of peanut (Arachis hypogaea L.) genotypes to smut (Thecaphora frezii) in the peanut growing region of Argentina. Peanut Sci. 48:61-67. doi: [: 10.3146/PS20-18.1].
Khoury C.K., Sotelo S.H., Hawtin G., Wibisono J., Amariles D., Guarino L., Kiene T. and Toledo A.. 2022. The plants that feed the world: baseline data and metrics to inform strategies for the conservation and use of plant genetic resources for food and agriculture. Item 16.2 of the Provisional Agenda Ninth Session of the Governing Body, 19–24 September 2022, New Delhi, India, 106 p.
Krapovickas A. and Gregory W.C.. 1994. Taxonomía del género Arachis (Leguminosae). Bonplandia. 8:1-186. doi: [: 10.30972/bon.81-43559].
Krapovickas A. and Gregory W.C.. 2007. Taxonomy of the genus Arachis (Leguminosae). Translated by D.E. Williams & C.E. Simpson. Bonplandia. 16(Supl.): 1-205. ISSN: 0524-0476.
Lazaridi E., Kapazoglou A., Gerakari M., Konstantina K., Passa K., Sarri E., Papasotiropoulos V., Tani E. and Babeli P.. 2024. Crop landraces and indigenous varieties: A valuable source of genes for plant breeding. Plants. 13(6):758. doi: [: 10.3390/plants13060758].
Leal-Bertioli S.C.M., Nascimento E.F.M.B., Chavarro M.C.F., Custodio A.R., Hopkins M.S., Moretzsohn M.C., Bertioli D. J. and Araujo A.C.G.. 2021. Spontaneous generation of diversity in Arachis neopolyploids (Arachis ipaënsis x Arachis duranensis)4x replays the early stages of peanut evolution. G3: Genes, Genomes, Genetics. 11(11): doi: 1 [: 0.1093/g3journal/jkab289].
Leal-Bertioli S.C.M., de Blas F.J., Chavarro M. C., Simpson C.E., Valls J.F.M., Tallury S.P., Moretzsohn M.C., Custodio A.R., Stalker H.T., Seijo G.J. and Bertioli D.J.. 2024. Relationships of the wild peanut species, section Arachis: A resource for botanical classification, crop improvement and germplasm management. Amer. J Bot. 111:e16357. doi: [: 10.1002/ajb2.1635].
Leal-Bertioli S.C.M., Hopkins M., Leverett J., Gonzales M., Tsai Y.C., Matusinec D., Tonnis B., Araujo A.C.G., and Bertioli D.J.. 2025. Registration of six disease resistant, high protein, induced allotetraploids derived from Arachis duranensi and A. ipaënsis, the genome progenitors of peanut. J. of Plant Reg. doi: [: 10.1002/plr2.70017].
Marden E., Hamilton R. S., Halewood M. and McCouch S.. 2023. International agreements and the plant genetics research community: A guide to practice. Proc. Natl. Acad. Sci. USA. 120 (14). doi: [: 10.1073/pnas.2205773119].
Massa A.N., Bressano M., Soave J.H., Buteler M.I., Seijo G., Sobolev V.S., Orner V.A., Oddino C., Soave S.J., Faustinelli P.C., de Blas F.J., lamb M.C. and Arias R.S.. 2021. Genotyping tools and resources to assess peanut germplasm: smut-resistant landraces as a case study. Peer J. 9:e10581. doi: [: 10.7717/peerj.10581].
Mosciaro M. J., Seghezzo L., Texeira M., Paruelo J. and Volante J.. 2023. Where did the forest go? Post-deforestation land use dynamics in the dry chaco region in Northwestern Argentina. Land use policy. 129. doi: [: 10.1016/j.landusepol.2023.106650].
Newman C.S., Andres R.J., Youngblood R.C., Campbell J.D., Simpson S.A., Cannon S.B., Scheffler B.E., Oakley A.T., Hulse-Kemp A.M. and Dunne J.C.. 2023. Initiation of genomics-assisted breeding in Virginia-type peanuts through the generation of a de novo reference genome and informative markers. Frontiers in Plant Sci. 13( 2022). doi: [: 10.3389/fpls.2022.1073542].
Pittman R.N. 1995. United States Peanut Descriptors. ARS-132, USDA-ARS.
Rago A. M., Cazón L. I., Paredes J.A., Molina J.P.E., Conforto E.C., Bisonard E.M. and Oddino C.. 2017. Peanut smut: from an emerging disease to an actual threat to Argentine peanut production. Plant Disease. 101:400-408.
. 2011. Nagoya protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilization to the convention on biological diversity: text and annex. Montreal, Canada, Secretariat of the Convention on Biodiversity, 15pp. doi: [: 10.25607/OBP-789].
Shahid M. A., Leaks K., Blount A. R. and Mackowiak C.. 2023. Perennial peanut as a potential living mulch and nitrogen source for citrus and other orchard crops in Florida. https://edis.ifas.ufl.edu/publication/HS1474.
Simpson C. E. 1984. Plant Exploration: planning, organization, and implementation with special emphasis on Arachis. pp. 1-20. In: Brown W. L., Chang T.T., Goodman M.M., and Jones Q. (eds.). Conservation of Crop Germplasm-An International Perspective. Crop Sci. Soc. of Amer. Madison, WI. USA.
Simpson C.E. and Starr J.L.. 2001. Registration of ‘COAN’ peanut. Crop Sci. 41:918.
Simpson C.E., Higgins D.L., Thomas G.D. and Howard E.R.. 1992. Catalog of passport data and minimum descriptors of Arachis hypogaea L. germplasm collected in South America 1977-1986. Texas Agric. Exp. Sta. MP-1737.
Simpson C.E., Starr J.L., Nelson S.C., Woodward K.E. and Smith O.D.. 1993. Registration of TxAG-6 and TxAG-7 peanut germplasm lines. Crop Sci. 33:1418.
Simpson C.E., Starr J.L., Church G.T., Burow M.D. and Paterson A.H.. 2003. Registration of ‘NemaTAM’ Peanut. Crop Sci. 43:1561.
Stalker H.T. 1993. Registration of four interspecific peanut germplasm lines resistant to Cercospora arachidicola. Crop Sci. 33:1117.
Stalker H. T. 2017. Utilizing wild species for peanut improvement. Crop Sci. 57:1102-1120.
Stalker H.T. and Simpson C. E.. 1995. Germplasm resources in Arachis. pp. 14-53. In: Pattee H.E. and Stalker H.T. (eds.), Advances in peanut science. APRES, Stillwater, OK.
Stalker H.T., Beute M.K., Shew B. B. and Barker K.R.. 2002a. Registration of two root-knot nematode-resistant peanut germplasm lines. Crop Sci. 42:312–313.
Stalker H.T., Beute M.K., Shew B. B. and Isleib T.G.. 2002b. Registration of five leaf spot-resistant peanut germplasm lines. Crop Sci. 42:314–316.
Stalker H.T., Tallury S., Ozias-Akins P., Bertioli D. and Leal-Bertioli S.C.. 2013. The value of diploid peanut relatives for breeding and genomics. Peanut Sci. 40: 70–88. doi: [: 10.3146/PS13-6.1].
Stalker H.T., Tallury S.P., Dunne J.C., Andres R.J., Hancock W.G. and Massa A.N.. 2025. Registration of two Arachis hypogaea x A. diogoi introgression lines. J. Plant Registrations. 19(3):e70018. doi: [: 10.1002/plr2.70018].
Swarup S., Cargill E.J., Crosby K., Flagel L., Kniskern J. and Glenn K.C.. 2021. Genetic variability is indispensable for plant breeding to improve crops. Crop Sci. 61:839-852.
Tallury S.P., Isleib T. G., Copeland S. C., Rosas-Anderson P., Balota M., Singh D. and Stalker H.T.. 2014. Registration of two multiple disease-resistant peanut germplasm lines derived from Arachis cardenasii Krapov. & W.C. Gregory, GKP 10017. J. Plant Reg. 8:86–89.
Tallury S.P., Tonnis B.D., Wang M.L., Mobley M.B., Brooks A., Stigura N. and Harrison M.L.. 2023. USDA peanut germplasm collection and its value for peanut improvement. Abstract. AAGB Conference, Huntsville, AL. October 16-19.
Tirnaz S., Zandberg J. , Thomas W.J.W., Marsh J., Edwards D. and Batley J.. 2022. Application of crop wild relatives in modern breeding: An overview of resources, experimental and computational methodologies. Frontiers in Plant Sci. 13 (2022). doi. [: 10.3389/fpls.2022.1008904].
Tyagi R. K. 2022. Networks and partnerships are tools for efficient PGR management. Indian J. Plant Genet. Resour. 35: 53–56.
. 1992. Convention on Biological Diversity. New York.
Upadhyaya H. D., Dwivedi S. L., Nadaf H.L. and Singh S.. 2011. Phenotypic variability and identification of wild Arachis accessions with useful agronomic and nutritional traits. Euphytica. 182:103-115.
Wang M. L., Barkley N. A., Chinnan M., Stalker H.T. and Pittman R.N.. 2010. Oil content and fatty acid composition variability in wild peanut species. Plant Genetic Resources. 8:232-234.
Wang M. L., Chen C. Y., Tonnis B., Barkley N.A., Pinnow D.L., Pittman R.N., Davis J., Holbrook C.C., Stalker H.T. and Pederson G.A.. 2013. Oil, fatty acid, flavonoid and resveratrol content variability and FAD2A functional SNP genotypes in the U.S. peanut mini-core collection. J. Agric. Food Chem. 61:2875-2882.
Wang M.L., Tonnis B., An Y., Pinnow D.L., Tishchenko V. and Pederson G.A.. 2015. Newly identified natural high-oleate mutant from Arachis hypogaea subsp. hypogaea. Mol. Breed. 35:186. doi: [: 10.1007/s11032-015-0377-3].
Wang M. L., Tonnis B.D., Chen C.Y., Li X., Pinnow D.L., Tallury S., Stigura N., Pederson G.A. and Harrison M.. 2022. Evaluation of variability in seed coat color, weight, oil content and fatty acid composition within the entire USDA-cultivated peanut germplasm collection. Crop Sci. 62:1-15.
Williams D. E. 2022. Global strategy for the conservation and use of peanut genetic resources. pp. 1-91 In Global Crop Diversity Trust. Bonn, Germany.
Williams K. and Williams D.. 2001. Evolving political issues affecting international exchange of Arachis genetic resources. Peanut Sci. 28:132-135. doi: [: 10.3146/i0095-3679-28-2-15].
Williams C., Walsh A., Vaglica V., Sirakaya A., da Silva M., Dalle G., Winterton D., Annecke W., Smith P., Kersey P.J., Way M., Antonelli A. and Cowell C.. 2020. Conservation Policy: Helping or hindering science to unlock properties of plants and fungi. Plants, People, Planet. 2:535–545. doi: [: 10.1002/ppp3.10139].
Yang C., Li Q., Wang X., Cui A., Chen J., Liu H., Ma W., Dong X., Shi T., Meng F., Yan X., Ding K. and Wu G.. 2023. Human expansion-induced biodiversity crisis over Asia from 2000 to 2020. Research. 6:0226. doi: [: 10.34133/research.0226].
Notes
- USDA-ARS, Griffin, GA [^]
- Department of Plant Pathology, University of Georgia, Athens, GA Institute of Plant Breeding, Genetics & Genomics, University of Georgia, Athens, GA [^]
- Department of Crop and Soil Sciences, University of Georgia, Athens, GA Institute of Plant Breeding, Genetics & Genomics, University of Georgia, Athens, GA [^]
- Texas A&M AgriLife Research, Stephenville, TX [^]
- FACENA-National University of Northeast, National Council for Scientific and Technological Research, IBONE (UNNE-CONICET), Corrientes, Argentina [^] Corresponding author (shyam.tallury@usda.gov)