

Introduction
Peanut smut is currently the most problematic disease for the Argentine peanut industry but also threatens peanut production worldwide through the movement of infected seed and contaminated goods. Thecaphora frezzii, the fungus causing peanut smut, was first discovered in 1962 on Arachis kulhmannii in Mato Grosso do Sul, Brazil (Carranza and Lindquist 1962, Krapovickas and Gregory 2007), but for decades, it was not found on cultivated peanut, A. hypogaea. However, in 1995, T. frezzii was discovered in a commercial production area in Argentina’s Córdoba province, and the fungus has since spread to all peanut-growing regions within the country (Rago et al. 2017, Paredes et al. 2024). Severely infested fields may have more than 50% disease incidence (Oddino et al., 2010, Bonessi et al. 2011, Marraro Acuña et al., 2013), and few effective chemical or practical cultural management strategies are available (Paredes et al., 2021). Since approximately 28M ha of peanut are planted worldwide (USDA-ERS 2020), T. frezzii has attracted international attention. Australia and South Africa currently have import restrictions on raw or unprocessed peanuts from Argentina, Bolivia, and Brazil (Australian Biosecurity Import Conditions 2017, Oilseeds Advisory Committee 2022).
Due to the lack of economical and effective chemical control measures, host resistance is the most promising approach for managing peanut smut. Conversely, widespread planting of the susceptible cultivar Granoleico likely dispersed the fungus throughout Argentina via infected seed and increased soilborne inoculum in previously infested areas. Granoleico constituted over 80% of Argentina’s peanut production between 2010 and 2018 (Paredes et al., 2024). Over the past five years, significant progress has been made in identifying resistant genotypes and genetic markers for resistance to T. frezzii, as well as in developing commercial cultivars resistant to peanut smut (Bressano et al., 2019, de Blas et al., 2019, 2021, Wann et al., 2020, Massa et al., 2021, Chamberlin et al., 2022, 2024). Host resistance appears to be controlled by one or two loci in cultivated peanut, A. hypogaea, simplifying breeding efforts (de Blas et al., 2021, Massa et al., 2021, Chamberlin et al., 2024). Since 2019, five Argentine cultivars with resistance to peanut smut have been released, but they are not widely planted due to low seed production and lack of commercial acceptance (Paredes et al., 2024). In the 2022-2023 season, the resistant cultivars accounted for only 8% of Argentina’s production (Paredes et al., 2024), and Granoleico was still planted on over 30% of the planted acres (SISA, 2023).
A major bottleneck in developing commercially desirable cultivars with smut resistance is the prevailing method of disease phenotyping, which is inefficient: opening individual pods by hand and looking for the presence of sori or teliospores (Rago et al., 2017, de Blas et al., 2019, Wann et al., 2020, Chamberlin et al., 2022). Opening pods by hand is necessary because signs of the fungus are only present in the pods of infected plants (Rago et al., 2017, Ospina-Maldonado et al., 2022), and a simple visual assessment for pod hypertrophy is insufficient. Not all diseased pods are deformed and assessing the presence or absence of hypertrophy is not always straightforward (Bennett et al., 2021). Once pods are opened, breeding programs rate disease incidence (proportion of pods with presence or absence of the T. frezzii teliospores) or severity. Disease severity is generally rated on an ordinal scale, such as that of Astiz Gassó et al., (2008): 0, pathogen not present; 1, seed with small/few sori; 2, partially damaged seed; 3, one kernel completely replaced by spores; and 4, all seeds completely replaced by spores. Because disease incidence (presence or absence of disease) is highly correlated with disease severity (Marraro Acuña et al., 2013, Bennett et al., 2021, Paredes et al., 2022), some labs measure disease incidence alone since it is considerably easier (Chamberlin et al., 2022). However, alternative rating approaches, such as counting only the proportion of severely damaged pods (3 and 4 on the Astiz Gassó scale), have been proposed (Paredes et al., 2022). If the number of genotypes being screened is high and labor is limited, disease ratings from one season may not be completed until seed for the next season is being planted, which slows down breeding efforts considerably.
Few alternatives to hand-opening pods have been explored. One method attempts to account for relative density: healthy peanut pods filled with seed are more dense than infected pods filled with T. frezzii teliospores (Bennett et al., 2021). Differences in density have been used to sort mycotoxin-contaminated wheat, corn, and peanut (Tkachuk et al., 1991, Rucker et al., 1994, Aoun et al., 2020, Stafstrom et al., 2021), as well as for assessing seed weight and maturity of in-shell peanuts (Sorensen et al., 2020, Domhoefer et al., 2022, Qiu et al., 2023). Capitalizing on the differences in density, Valente and colleagues (2014, 2017) were able to distinguish between infected and healthy pods, as well as estimate the percentage of seeds that were damaged using an experimental X-ray microtomography system. Unfortunately, this X-ray system is not commercially available. Lewis et al., (2024) recently evaluated the potential of a resonant cavity instrument for phenotyping peanut smut, but this instrument is also experimental and commercially unavailable. Based on the positive results from the experimental X-ray microtomography system, other digital X-ray technologies were deemed to be possible solutions. The objective of this project was to construct and evaluate a simpler and more economical (<$25,000) X-ray imaging system for phenotyping resistance to peanut smut. The system was evaluated by having human raters visually assess images of healthy and faux-infected peanut pods taken by this X-ray system. Results from a preliminary dataset were presented recently (Bennett et al., 2024).
Materials and Methods
X-ray Imaging System.
An X-ray digital imaging system for peanut pods was designed and constructed from the following off-the-shelf components (Figure 1): an X-ray tube source (Jupiter 5000, Oxford Instruments, Scotts Valley, CA); a line-scan X-ray camera (Harrier X-Ray CMOS TDI Line-Scan Detector, X-Scan Imaging, San Jose, CA); and a sample conveyor system driven by a stepper motor (FSL40E25010C7, FUYU Technology Co. Ltd., Sichuan, China). The X-ray tube control box and power unit were attached to an enclosure made from lead-lined metal and a single X-ray-shielded lead glass viewing window. The metal enclosure walls were built using lead and aluminium sheets, both with 4-mm thickness, to minimize X-ray leakage. The bottom of the enclosure was in the direct path of the X-ray beams, so two layers of lead sheets were used for its construction. Measured data showed zero radiation above background levels in all locations next to the enclosure. This feature simplifies compliance with radiation safety regulations.
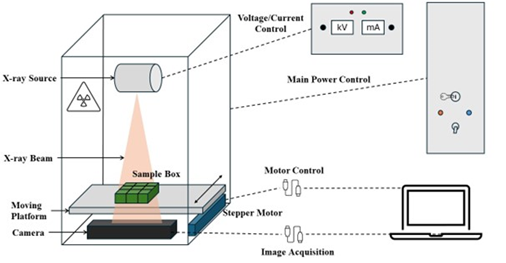
Figure 1. Schematic diagram of the X-ray imaging system.
The X-ray tube light source, mounted 17 cm above the sample-carrying conveyor, generated a 15.2-cm-diameter diverging cone-shaped X-ray beam on the sensing area of the line-scan camera (Figure 1). The sample-carrying conveyor was installed above the camera, and an 18.5 x 15.5 x 4.3-cm polypropylene craft organizer box (Vidifor through Amazon, Seattle, WA) was used to move nine pods at a time through the camera’s field of view. Removable dividers in the box were installed to make nine 5.8 x 4.8 x 3.8-cm grids, which accommodated the range of pod sizes found in runner, Spanish, Virginia, and Valencia peanut market types. A power unit was built to house a 24V/100W power adaptor for the X-ray imaging system (Figure 2). To meet Oklahoma State University’s Radiation Safety Office requirements, as well as federal and state regulations, several safety switches and warning lights were installed outside of the power unit. An enclosure was also constructed to house the controls for the X-ray tube power settings and the movement of the sample-carrying conveyor.

Figure 2. Exterior view of the X-ray imaging system (a); interior view with (A) X-ray source, (B) sample conveyer platform, and (C) X-ray camera (b); power unit (c); and control box (d).
Phantom Test Pods.
Pods from four cultivars representing each of the four major market types were used to test the X-ray system: Southwest Runner (Kirby et al., 1998), OLé (Spanish; Chamberlin et al., 2015), Valencia C (Hsi 1980), and Jupiter (Virginia). In medical imaging, test phantoms are objects used to simulate human tissue and organs (McGarry et al., 2020). Since T. frezzii is not present in the U.S., suitable test phantoms were made as a proxy for infected pods using plant-based powders or fungal spores. Faux-infected pods were made by carefully opening the pericarp along the suture and replacing all or some seeds with ground coffee, corn starch, or teliospores of the corn smut fungus, Mycosarcoma maydis. The corn smut sori were collected in 2022 from Oklahoma State University’s Caddo Research Station in Fort Cobb. A thin strip of Elmer’s Liquid School Glue (Newell Brands, Atlanta, GA) was applied along the rim of the suture to reattach the pericarp halves. The pod was held together until the glue dried by placing them between the rods of an Endicott-Seymour (Ann Arbor, MI) drying rack.
A total of nine phantom pod types were made (Table 1). The three “healthy” pod types, equivalent to the Astiz Gassó et al., (2008) severity rating of “0”, included the following: 1) normal, where the pod pericarp/shell was opened and reglued; 2) empty, pods with all seeds removed; and 3) part-seed, pods with at least one seed removed. The empty and part-seed pods were included to mimic “pops”, i.e. unfilled pods (Brady et al., 1948). Faux-infected “unhealthy” pod types included: 4) full coffee, all seeds replaced by finely ground coffee; 5) part coffee, one seed replaced by coffee; 6) full corn starch, all seeds replaced by cornstarch; 7) part corn starch, one seed replaced by cornstarch; 8) part corn smut, one seed replaced by corn smut teliospores; and 9) one seed with a faux peanut smut sorus. Disease severity levels of “4” and “3” (Astiz Gassó et al., 2008) were made by filling entire pods or individual locules with coffee, corn starch, or corn smut teliospores by pressing the materials into the pods until they were level with the suture, placing a thin strip of glue along the suture, and quickly bringing both halves together. The faux sori were made to approximate disease severity levels 1 or 2 of Astiz Gassó (2008). To make the faux sori (Figure 3), a cavity approximately 0.5 cm in diameter and 0.25-cm deep was made on the surface of one seed with a dissecting needle. A faux peanut smut “sorus” was made by rolling ca. 0.5-cm3 of M. maydis teliospores in a thin sheet of Parafilm (Amcor, Neenah, WI). The roll was cut to size, twisted to seal open ends, shaped to fit in the cavity, and attached to the seed with glue. A total of 3,303 pods were used to evaluate the X-ray system.
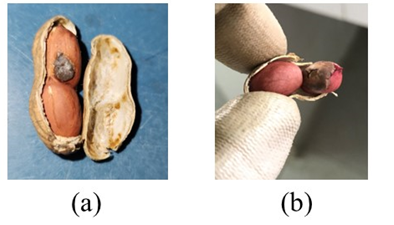
Figure 3. Faux peanut smut “sorus” made with corn smut teliospores (a), and a Thecaphora frezzii sorus (b).

Number of “healthy” and faux-infected phantom pods used to evaluate the X-ray system for each market type.
X-ray Imaging and Rating of Pods.
A motor-driven conveyor was used to carry a sample box with nine grids to the field of view of the X-ray camera. Gray-scale X-ray images of pods were captured line by line by the line-scan X-ray camera (X-88, X-Scan Imaging, San Jose, CA). The X-ray tube was set at 0.5mA and 30kV. A graphical user interface (GUI) on the laptop was developed to control the overall image acquisition process. All images were automatically stored in a dedicated folder. Since each scanned image contained nine pods, an Open Source Computer Vision Library (OpenCV.org) script was made to automatically segment single pod images using the fixed coordinates of the grids in the craft organizer box. The script also added a file name to each segmented image.
Images from each of the phantom pod types and market types were divided into three groups (blocks) and uploaded to a dedicated Google Drive. Image names and file locations (URLs) were placed in a Google Sheet by block, and each image was assigned a random number using the RAND() function and sorted numerically. The four market types/cultivars were rated together. Online scoring sheets were created on Google Forms, and each block was divided into five sets of approximately 220 images to reduce rater fatigue. Raters were asked to review three to four training images of each of the three healthy and six unhealthy phantom pod types (Figure 4) before rating each image as healthy or unhealthy. Google Forms was used to export the ratings into a single spreadsheet.
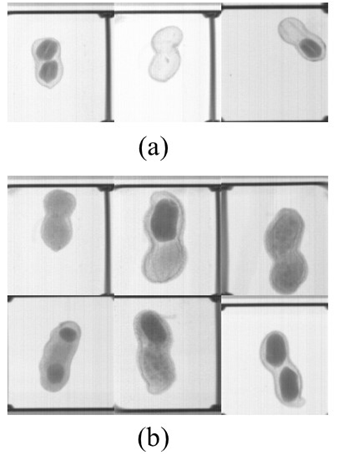
Figure 4. Sample images of the phantom test pod types used as part of the training set for raters. “Healthy” pods (a), and “Unhealthy” pods filled with ground coffee, corn starch, corn smut teliospores, or an artificial corn smut “sorus” (b).
Experimental Design and Data Analysis.
As described previously, the images were organized into a randomized complete block design with three blocks. Chi square tests (PROC FREQ, SAS Version 9.4) were used to examine rater accuracy within cultivars/market type and phantom pod type. Differences in accuracy among raters and over time were also examined.
Results and Discussion
Overall, raters correctly identified “healthy” and “unhealthy” phantom test pods using the X-ray images (Table 2) with some accuracy (85%). However, some pod types were easier to score than others (P < 0.01). Healthy pods were easier to identify (89.5%) than faux-infected pods (82.6%). Most notably, raters had the greatest difficulty in correctly scoring pods with the artificial sorus (21.2% correct). Raters were also more likely to score Valencia pods incorrectly, but this is likely a result of having twice as many Valencia pods with the artificial sorus (Table 1). As our limited quantity of corn smut teliospores was depleted, we focused on more pods with the artificial sorus since this pod type required fewer teliospores and was likely to be most difficult to detect. All ten raters were able to score at least 80% of the images correctly, but there was a significant, ~7% difference between the most and least accurate rater (P <0.01). On the other hand, raters might improve slightly with practice (P <0.01). More images in the third and last block of images were scored correctly than in the first two blocks.

Success of 10 raters in scoring X-ray images of phantom test pods as healthy or unhealthy.
These results indicate that the X-ray system has potential as a more efficient method for phenotyping peanut smut than hand-opening pods; however, it needs to be tested in Argentina on pods that are healthy and pods that are infected with varying severity levels of peanut smut. Lacking easy access to a biosafety level 3 (BSL-3) lab with radiation safety capabilities, we used faux-infected phantom test pods to evaluate the X-ray system. Corn starch, ground coffee—even M. maydis teliospores—may not be accurate test phantoms for visualizing T. frezzii under X-ray. Unlike the single spore teliospores of M. maydis, T. frezzii teliospores are rarely single-spored and more often tightly fused into balls composed of 2-10 spores. Individual teliospores of T. frezzii are also larger, 15-22 x 16-22 um vs. 7-11 x 7-13 um for M. maydis (Vánky, 2012). In addition, previous work showed that the density of pods infected with T. frezzii was approximately 437.5 g/L for mature pods and 380.5 g/L for immature pods (Bennett et al., 2021). Initially, we tried to fill pod locules at a specific density, but the process required estimating pod cavity volume—a task too laborious for the large number of pods needed to test the system. As a result, the density of our faux-infected phantom test pods may differ from that of pods infected with peanut smut.
Another limitation of the current X-ray system setup was the inability to generate images from which raters could consistently distinguish pods with the artificial corn smut sorus. Fortunately, pods with disease severity levels of 1 (seed with small/few sori) and 2 (partially damaged seed) appear to be rare in both smut-susceptible and -resistant peanut genotypes (Paredes et al., 2022). We are currently testing if better images can be obtained using different exposure times and X-ray intensities. While the image resolution of this system is less than the experimental X-ray microtomography system tested by Valente and colleagues (2017), this system is considerably more affordable. The cost for materials (light source, camera, lead sheets, computer, etc.) was less than $25,000. To increase the efficiency of this system, we are also testing machine learning algorithms for automated scoring. While the X-ray system scanned nine pods in 15 sec, additional time was needed to fill and empty the sample box, operate the system, and score images. For genotypes with smaller pods, like Spanish and small-seeded runners, efficiency could be increased further by placing more non-overlapping pods in each grid.
Recent discoveries of molecular markers associated with resistance to peanut smut greatly help peanut breeders develop improved cultivars (Massa et al., 2021, Chamberlin et al., 2024). Despite these advances, the need for more efficient alternatives to hand-opening pods will likely continue. Not all countries may have enacted as stringent measures to prevent the introduction of infected seeds as Australia and South Africa have (Australian Biosecurity Import Conditions 2017, Oilseeds Advisory Committee 2022), and the fungus may move to new production areas. To date, T. frezzii has been reported from Bolivia (Soave et al., 2014) and Brazil (Carranza and Lindquist 1962) in addition to Argentina. In recent decades, agriculture has steadily moved into wild ecosystems, and large-scale peanut production is now present in the Gran Chaco region of northern Argentina and Paraguay (USDA-Foreign Agricultural Service). These areas are documented to have at least nine wild peanut species (Krapovickas and Gregory 2007, Morales et al., 2019), and peanut monocultures in these areas increase opportunities for encountering new genotypes of T. frezzii from wild peanuts that may be able to overcome the currently available resistance. Much work remains in developing an efficient phenotyping method for peanut smut, but the X-ray system presented here is one starting point.
Acknowledgements
The authors thank: Angie Harting, Paula Lor, Jorin Stockebrand, Austin Pickering, Jerry J. Wang, Joe Preston, Mike Fleming, Nick Semtner, and Mike Veldman for technical assistance; Michelle Allen, Andrew Hahn, and Elisabeth Tidwell for work on an X-ray system prototype; and Iris Bennett, Kyren Bunyard, Dianne Koch, Chijioke Nkwocha, Clint Slocum, Lohini Suresh, and Karen Weckler for helping to rate pods. This research was supported by USDA-ARS CRIS Project No. 3072-21220-009-00D, USDA-ARS National Plant Disease Recovery System, and MARS, Inc. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Literature Cited
Aoun M., Stafstrom W., Priest P., Fuchs J., Windham G.L.,Williams W.P., and Nelson R.J. 2020. Low-cost grain sorting technologies to reduce mycotoxin contamination in maize and groundnut. Food Control. 118:107363. doi: [: 10.1016/j.foodcont.2020.107363].
Astiz Gassó M.M., Leis R., and Marinell A. 2008. Evaluación de incidencia y severidad del carbón de maní (Thecaphora frezii) en infecciones artificiales, sobre cultivares comerciales de maní. 1° Congreso Argentino de Fitopatología. Córdoba, Argentina. Page 118 [: (abst.)].
Australian Biosecurity Import Conditions. 2017. BICON - Case Alert. https://bicon.agriculture.gov.au/BiconWeb4.0/ViewElement/Element/Alert?elementPk=687363&casePk=682337 2024September7.
Bennett R.S., Rodriguez A.V., Baldessari J.J., Chamberlin K.D., Payton M.E., and Wang N. 2021. A note on the association between Thecaphora frezzii infection and peanut pod density. Peanut Sci. 48:54–60. doi: [: 10.3146/PS20-27.1].
Bennett R.S., Wang N., Weckler P.R., Zhang Y., Payton M.E., Baldesarri J., and Chamberlin K.D. 2024. Towards efficient phenotyping for peanut smut resistance: line-scan X-ray imaging of faux-infected pods. 2024 Proc. Amer. Peanut Res. Educ. Soc. 56:156 [: (abstr.)].
Bonessi F., Rago A., Marinelli A., March G., Cazón I., Garcia J., arditi L., D’Eramo L., Ferrari S., y Oddino C. 2011. Efecto de la fertilización sobre la intensidad del carbón del maní. XXVI Jornada Nacional de Maní, General Cabrera, Córdoba, Argentina.
Brady N.C., Reed J.R., and Colwell W.E. 1948. The effect of certain mineral elements on peanut fruit filling. Agron. J. 40:155–167.
Bressano M., Massa A.N., Arias R.S., de Blas F., Oddino C., Faustinelli P.C., Soave S., Soave J.H.,Pérez M.A., Sobolev V.S., Lamb M.C., Balzarini M., Buteler M.I., and Seijo J.G. 2019. Introgression of peanut smut resistance from landraces to elite peanut cultivars (Arachis hypogaea L.). PLOS One. 14: [: e0211920].
Carranza J.M., and Lindquist J.C. 1962. Thecaphora frezii n. sp., parásita de Arachis sp. Bol. Soc. Argent. Bot. 10:11–18.
Chamberlin K.D., Baldessari J., Bennett R.S., Clevenger J.P., Holbrook C.C., Tallury S.P., Chu Y., Ozias-Akins P., Conde M.B., and Payton M.E. 2022. Identification of germplasm resistant to peanut smut. Peanut Sci. 49:1–16. doi: [: 10.3146/0095-3679-491-PS21-10].
Chamberlin K.D., Bennett R.S., Baldessari J., De G. La Barrera, Cordes G., Grandon N.G., Mamani E.M., Rodriguez A., Morchetti S., Holbrook C.C., Ozias-Akins P., Chu Y., Tallury S., Clevenger J., Korani W., Scheffler B., Youngblood R.C., and Simpson S. 2024. Discovery of a resistance gene cluster associated with smut resistance in peanut. Peanut Sci. 51:59–65. doi: [: 10.3146/0095-3679-51-PS23-6].
Chamberlin K.D., Bennett R.S., Damicone J.P., Godsey C.B., Melouk H.A., and Keim K. 2015. Registration of ‘OLé’ peanut. J. Plant Regist. 9:154–158.
Domhoefer M., Chakraborty D., Hufnagel E., Claußen J., Wörlein N., Voorhaar M., Anbazhagan K., Choudhary S., Pasupuleti J., Baddam R., Kholova J. and Gerth S. 2022. X-ray driven peanut trait estimation: computer vision aided agri-system transformation. Plant Methods. 18:1–14.
Hsi D.C. 1980. Registration of New Mexico Valencia C peanut (Reg. No. 24). Crop Sci. 20:113–114. doi: [: 10.2135/cropsci1980.0011183X002000010033x].
Kirby J.S., Melouk H.A., Stevens T.E., Banks D.J., Sholar J.R., Damicone J.P., and Jackson K.E. 1998. Registration of ‘Southwest Runner’ peanut. Crop Sci. 38:545–546. doi: [: 10.2135/cropsci1980.0011183X002000010033x].
Krapovickas A., and Gregory W.C. 2007. Taxonomy of the genus Arachis (Leguminosae). Translated by D.E. Williams & C.E. Simpson. Bonplandia. 16 (Supl.):1–205.
Lewis M.A., Trabelsi S., Bennett R.S., and Chamberlin K.D. 2024. Utilization of a resonant cavity for characterization of single in-shell peanuts. Food Analytical Methods. 17:855–866.
Marraro Acuña F., Cosa M.T., and Wiemer A.P. 2013. Carbón del maní: histopatología, incidencia y severidad. XXVIII Jornada Nacional de Maní. General Cabrera, Córdoba, Argentina.
Massa A.N., Bressano M., Soave J.H., Buteler M.I., Seijo G.,Sobolev V.S., Orner V.A., Oddino C., Soave S.J., Faustinelli P.C., de Blas F.J., Lamb M.C., and Arias R.S. 2021. Genotyping tools and resources to assess peanut germplasm: smut-resistant landraces as a case study. PeerJ. 9:e10581. doi. [: 10.7717/peerj.10581].
McGarry C.K., Grattan L.J., Ivory A.M., Leek F., Liney G.P., Liu Y., Miloro P., Rai R., Robinson A.P., Shih A.J., Zeqiri B., and Clark C.H. 2020. Tissue mimicking materials for imaging and therapy phantoms: a review. Phys. Med. Biol. 65:23TR01. doi: [: 10.1088/1361-6560/abbd17].
Morales M., Oakley L., Sartori A.L.B., Mogni V.Y., Atahuachi M., Vanni R. O., Fortunato R.H., and Prado D.E. 2019. Diversity and conservation of legumes in the Gran Chaco and biogeograpical inferences. PLOS One. 14:e0220151.
Oddino C., Marinelli A., March G.J., Tarditi L., D’Eramo L., y Ferrari S. 2010. Relación entre el potencial inóoculo de Thecaphora frezii, la intesidad de carbón del maní y el rendimiento del cultivo. XXV Jornada Nacional del Maní. General Cabrera, Córdoba, Argentina.
Oilseeds Advisory Committee. 2022. Raw or unprocessed peanuts. Oil and Protein Seeds Development Trust (OPDT) / Oilseeds Advisory Committee (OAC). https://www.opot.co.za/index.php?page=all-countries-excluding-argentina-brazil-bolivia (accessed 8 September 2024).
Ospina-Maldonado S., Castlebury L.A., Bennett R.S., and Salgado-Salazar C. 2022. Peanut smut: a diagnostic guide. Plant Health Progress. 23:492–496.
Paredes J.A., Cazón L.I., Conforto E.C., and Rago A.M. 2024. Peanut smut in Argentina: an analysis of the disease, advances and challenges. Plant Dis. 108:2593-2606.
Paredes J.A., Cazón L.I., Oddino C., Monguillot J.H., Rago A.M., and Edwards Molina J.P. 2021. Efficacy of fungicides against peanut smut in Argentina. Crop Prot. 140:105403.
Paredes J.A., Edwards Molina J.P., Cazón L.I., Asinari F., Monguillot J.H., Morichetti S.A., Rago A.M., and Torres A.M. 2022. Relationship between incidence and severity of peanut smut and its regional distribution in the main growing region of Argentina. Trop. Plant Pathol. 47:233–244.
Qiu G., Liu Y., Wang N., Bennett R.S., and Weckler P.R. 2023. Non-destructive method for estimating seed weights from intact peanut pods using soft X-ray imaging. Agronomy. 13:1127.
Rago A.M., Cazón L.I., Paredes J.A., Molina J.P.E., Conforto E.C., Bisonard E.M., and Oddino C. 2017. Peanut smut: from an emerging disease to an actual threat to Argentine peanut production. Plant Dis. 101:400–408.
Rucker K.S., Kvien C.K., Calhoun K., Henning R.J., Koehler P.E., Ghate S.R., and Holbrook C.C. 1994. Sorting peanuts by pod density to improve quality and kernel maturity distribution and to reduce aflatoxin. Peanut Sci. 21:147–152. doi: [: 10.3146/i0095-3679-21-2-17].
Sistema de Información Simplificado Agrícola (SISA). 2023. Maní 2022 - 2023. Instituto Nacional de Semillas (INASE). 2024September9 https://www.argentina.gob.ar/sites/default/files/sisa_mani_22_23_v1.pdf.
Soave J., Bianco C., Burgoa R., Montaño R., Rago A., et al., 2014. Primera detección de carbón del maní (Thecaphora frezii) en Bolivia. 3° Congreso Argentino de Fitopatología. Asociación Argentina de Fitopatólogos, San Miguel de Tucumán, Argentina. Page 211.
Sorensen R.B., Butts C.L., and Lamb M.C. 2020. X-ray technology to determine peanut maturity. Peanut Sci. 47:38–45. doi: [: 10.3146/PS20-2.1].
Stafstrom W., Wushensky J., Fuchs J., Xu W., Ezera N., and Nelson R.J. 2021. Validation and Application of a Low-Cost Sorting Device for Fumonisin Reduction in Maize. Toxins. 13:652.
Tkachuk R., Dexter J.E., Tipples K.H., and Nowicki W. 1991. Removal by specific gravity table of tombstone kernels and associated trichothecenes from wheat infected with Fusarium head blight. Cereal Chem. 68:428–431.
USDA-Foreign Agricultural Service. 2024. Peanut Explorer. 2024September16 https://ipad.fas.usda.gov/cropexplorer/cropview/commodityView.aspx?cropid=2221000&sel_year=2024&startrow=1.
Valente M., Malano F.M., Perez P., and Baldessari J. 2014. Rayos X detectan y distinguen carbón, cascara y semillas en vainas intactas de maní. XXIX Jornada Nacional del Maní. General Cabrera, Córdoba, Argentina.
Valente M., Malano F.M., Perez P., and Baldessari J. 2017. Automated peanut smut damage assessment on intact pod bulks by using X-ray devices and proprietary software. Córdoba, Argentina.
Vánky K. 2012. Smut fungi of the world. American Phytopathological SocietySt. Paul, MN.
Wann D., Falco A., Cavigliasso M., and Cassano C. 2020. Phenotypic variation of peanut smut (Thecaphora frezii) incidence and severity in the US peanut mini-core collection. Peanut Sci. 47:46–53. doi: [: 10.3146/PS20-4.1].
Notes
- Department of Biosystems and Agricultural Engineering, Oklahoma State University, Stillwater, OK 74078 [^]
- Department of Biomedical Sciences, Rocky Vista University, Englewood, CO 80112 [^]
- Instituto Nacional de Tecnología Agropecuaria, Estación Experimental Agropecuaria Manfredi, Córdoba Province, Argentina [^]
- Peanut and Small Grains Research Unit, Oklahoma and Central Plains Agricultural Research Center, USDA-ARS, Stillwater, OK 74075 [^]
- Foreign Disease-Weed Science Research Unit, USDA-ARS, Fort Detrick, MD 21702 [^]
- Fundación para el Estudio de Especies Invasivas, Hurlingham, Buenos Aires Province, Argentina [^] Corresponding author’s E-mail: rebecca.bennett@usda.gov