

INTRODUCTION
Normal oleic (O) to linoleic (L) fatty acid ratios range from 1 to 3:1 for commercially available peanut (Arachis hypogaea L.) seed (Branch et al., 1990; Branch et al., 2020). However, high-oleic O/L ratios varies between 20 to 40:1; whereas, very high O/L ratios were usually >40:1 in south Georgia (Branch et al., 2020).
The importance of high-O/L ratios is to extend the shelf-life of peanut and peanut products (Mozingo et al., 2004). Unfortunately, high-oleic cultivars also result in extremely low-linoleic fatty acid content of approximately 2 to 3% compared to 20 to 30% for normal-oleic genotypes. Linoleic fatty acid (C18:2) is an essential fatty acid in humans and animals that must be supplied in the diet from plant sources like peanut and many other oilseed crops (Gurr and James, 1975).
Recently, Marklund et al., (2019) stated that current trends to increase use of high-oleic and low-linoleic fatty acids in several oilseed crops could increase the risk of insufficient polyunsaturated fatty acids (PUFA) consumption needed to prevent cardiovascular disease (CVD). Obviously, many other factors contribute to our overall health. Genetics, diets, and environmental factors each play a major role (CAST, 1991).
In the past, the term “mid-oleic” was used for peanut seed with an O/L ratio between 3 to 5:1. A few such cultivars, ‘NC 7’ (Wynne, et al., 1979) and ‘Virugard’ (released by the private breeding program of AgraTech, Inc.) were found to have slightly higher than normal-oleic O/L ratios (Branch, et al., 2003). Recently, the University of Georgia (UGA) peanut breeding program also developed a mid-oleic advanced breeding line, GA 132504.
In addition to genotypic differences in O/L ratios, environmental factors such as temperature and maturity interact with genotypes to influence O/L ratios. Warmer U.S. locations and mature seed result in higher O/L ratios (Ahmed and Young, 1982). Consequently, it is important to evaluate genotypes for O/L ratios at warm test locations. In south Georgia, O/L ratios between mid-oleic and high-oleic have predominantly been found to be heterozygous in segregating cross-populations, and such genotypes did not breed true-to-type. So, the objective of the genetic study was to determine the inheritance of mature mid-oleic peanut seed at the UGA, Tifton campus location in south Georgia.
Materials and Methods
Crosses were made in the greenhouse during the fall and winter months of 2017-18 between two mid-oleic genotypes, NC 7 and GA 132504, x ‘Tifguard’ (Holbrook et al., 2008) a normal-oleic cultivar and ‘TifNV-High O/L’ (Holbrook et al., 2017) a high-oleic cultivar that resulted from three backcrosses to Tifguard. In 2018, both the F1 seed and the four aforementioned parents were space-planted together approximately 122-cm apart in one-row plots, 6.1 m long by 1.8 m wide at the UGA Coastal Plain Experiment Station, Gibb’s Research Farm, Tifton, GA. The following year (2019), F2 seed from each cross combination were space-planted approximately 30-cm apart in two-row plots, 36.5 m long by 1.8 m wide.
These field nursery plots were in a three-year rotation following corn (Zea mays L.) and cotton (Gossypium sp.). Each year, the soil-type was a Tifton loamy sand (fine-loamy, siliceous, thermic, Plinthic Kandidult). University of Georgia Cooperative Extension Service recommended cultural practices with irrigation were used throughout each growing season.
Individual plants were harvested near optimum maturity based upon number of days after planting and above-ground appearance. After harvest, peanut pods were dried with forced warm-air to approximately 6% seed moisture content before weighing and shelling. Ten sound mature kernels (SMK) per plant were sent to W. M. Keck Metabolomics Research Lab at Iowa State University for oleic and linoleic fatty acid methyl ester (FAME) determination by GC from parents, F1, and F2 cross populations. After cross-pollination, the resulting seed (embryo and cotyledons) will be F1 hybrid tissue, while the testa and the whole female plant would still be maternal tissue (F0). Likewise, hybrid seed from individual F1 plants would produce F2 seed (embryo and cotyledons). Consequently for determining fatty acid profiles, the average of ten seed per individual plant were used to measure O/L ratio for each filial generation.
Data from percentages of oleic and linoleic fatty acid and O/L ratios were subjected to analysis of variance (ANOVA). Waller-Duncan Bayesian T-test (k-ratio = 100) was used for mean separation involving three or more entries; whereas, only a Student’s t-test was used for two mean separation by SAS (SAS, Cary, NC). Segregation data was analyzed by chi-square analysis for goodness-of-fit (P≤0.05) to expected genetic ratios (Strickberger, 1968).
Results and Discussion
Significant differences (P≤0.05) were found among percentages of oleic and linoleic fatty acids found in the peanut seed of the mid-oleic breeding line, GA 132504, and four normal-oleic runner-type cultivars (Table 1). The one-year (2015) and the two-year (4-tests) average showed that the mid-oleic breeding line had a significantly higher percent oleic and a significantly lower percent linoleic compared to the normal-oleic check cultivars. The data was equally consistent across tests and years. Average oleic to linoleic (O/L) fatty acid ratio (4.1:1) of the advanced Georgia breeding line was also found to be significantly higher than ‘Georgia-06G’ (Branch, 2007) over two years and four tests. In one test conducted during 2015, GA 132504 was again found to have a significantly higher O/L ratio (5.0:1) than four other normal-oleic cultivars. These data clearly show that GA 132504 is a mid-oleic with O/L ratios ranging from 3 to 5:1 in south Georgia (Table 1).

Four-Tests Average Seed Percentage of Oleic (O), Linoleic (L) Fatty Acid, and O/L Ratio for Four Normal-Oleic versus One Mid-Oleic (GA 132504) Peanut Genotypes when Grown under Maximum Input Production Practices with Irrigation at Tifton, GA 2014-15.
Cross combinations between mid-oleic NC 7 x normal-oleic Tifguard and mid-oleic GA 132504 x normal-oleic Tifguard had an average F1 plant (F2 seed) O/L ratio of 3.3:1 and 2.5:1, respectively (Fig. 1). Significant differences were found among parents and the F1 plants (F2 seed). However, F2 plant (F3 seed) segregation from these same two cross combinations did not show any high-oleic plants which strongly indicate that normal-oleic and mid-oleic are very similar with regard to alleles for the high-oleic trait (Table 2).
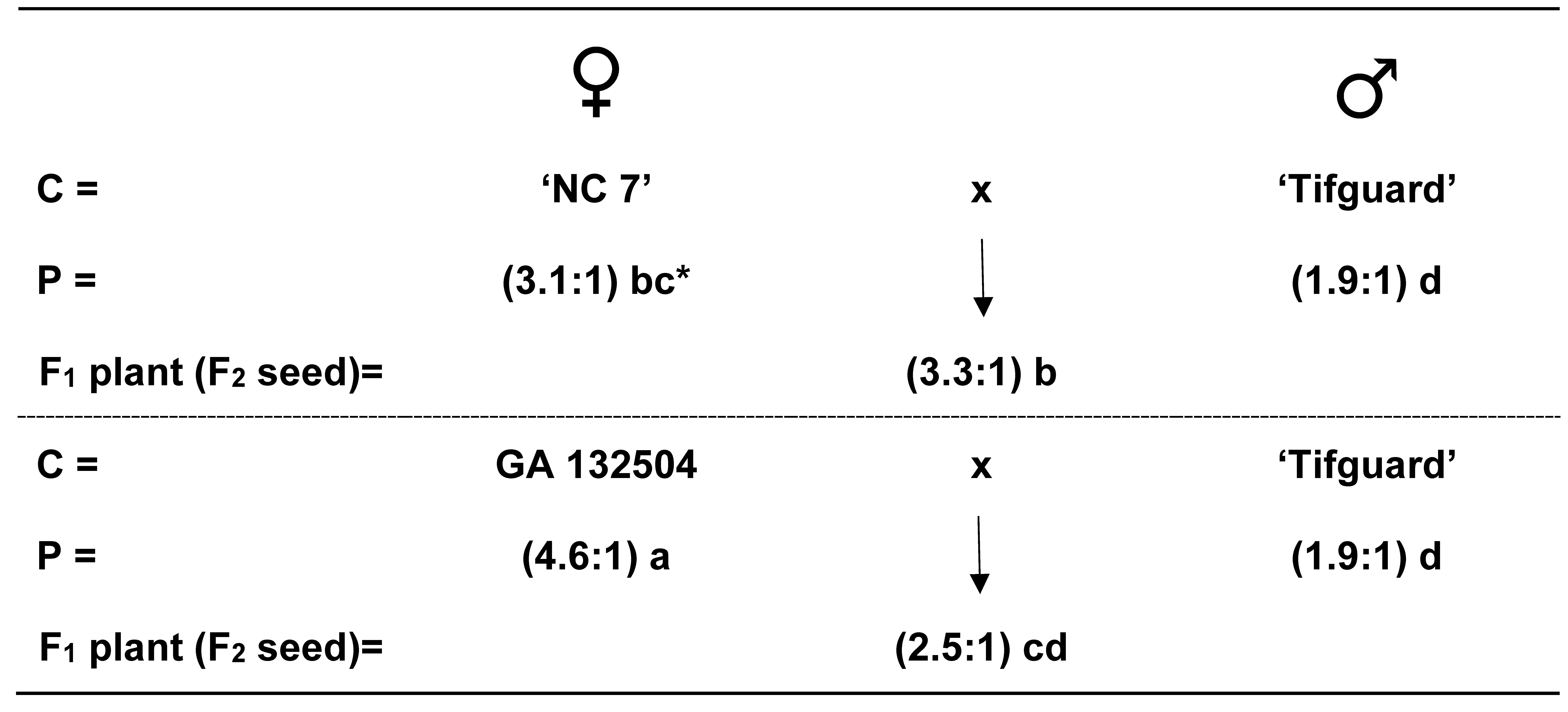
Figure 1. Average oleic to linoleic (O/L) fatty acid ratios from two cross combinations involving mid-oleic ‘NC 7’ x normal-oleic ‘Tifguard’ and mid-oleic GA 132504 x normal-oleic ‘Tifguard’. *Mean O/L ratios followed by the same letter not significantly different at P≤0.05.
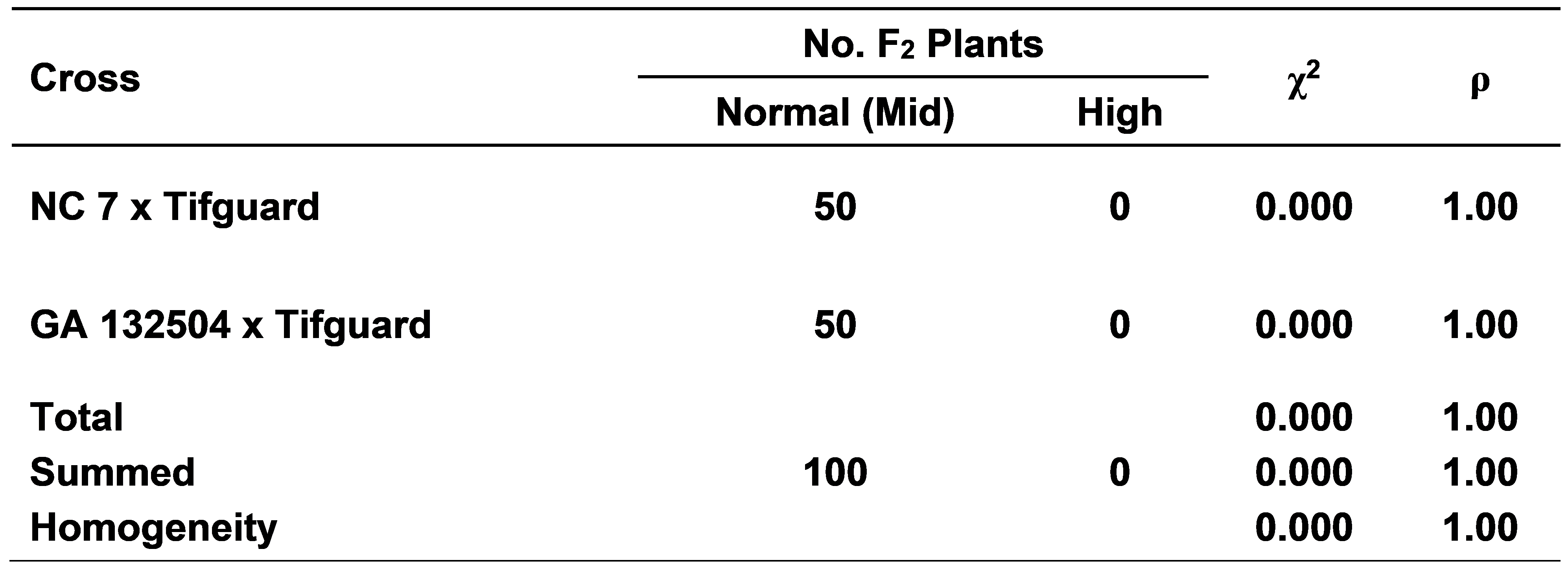
F2 individual plant (F3 seed) segregation for normal (mid) and high-oleic fatty acid ratios from two cross combinations.
In the mid-oleic x high-oleic cross combinations between NC 7 x TifNV-High O/L and GA 132504 x TifNV-High O/L resulted in an average F1 plant (F2 seed) O/L ratios of 8.1:1 and 9.0:1, respectively (Fig. 2). As expected, significant differences were found between the normal (mid)-oleic and high-oleic parents and hybrid seed. Likewise, the F2 plant (F3 seed) segregation from these same two cross combinations did show a similar acceptable fit for a 3 normal (mid)-oleic:1 high-oleic ratio (Table 3). Total, summed, and homogeneity chi-square values were also found acceptable for the 3:1 expected genetic ratio. These results agree with an earlier report by Chu et al., (2011) regarding a one-gene model for the high-oleic trait in TifNV-High O/L, then referred to as ‘Tifguard-High O/L’ since Tifguard was the recurrent parent.
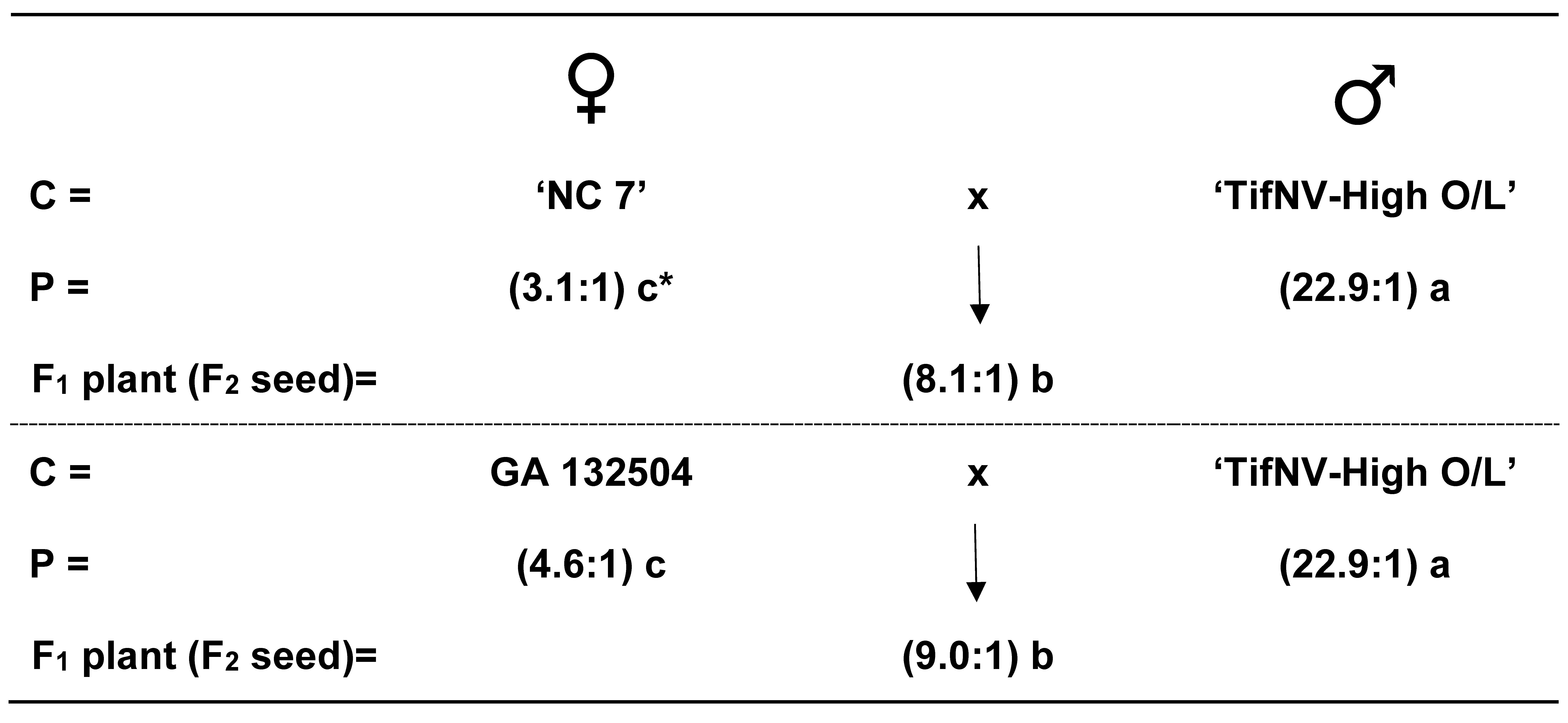
Figure 2. Average oleic to linoleic (O/L) fatty acid ratios from two cross combinations involving mid-oleic ‘NC 7’ x high-oleic ‘TifNV-High O/L’ and mid-oleic GA 132504 x high-oleic ‘TifNV-High O/L’. *Mean O/L ratios followed by the same letter not significantly different at P≤0.05.
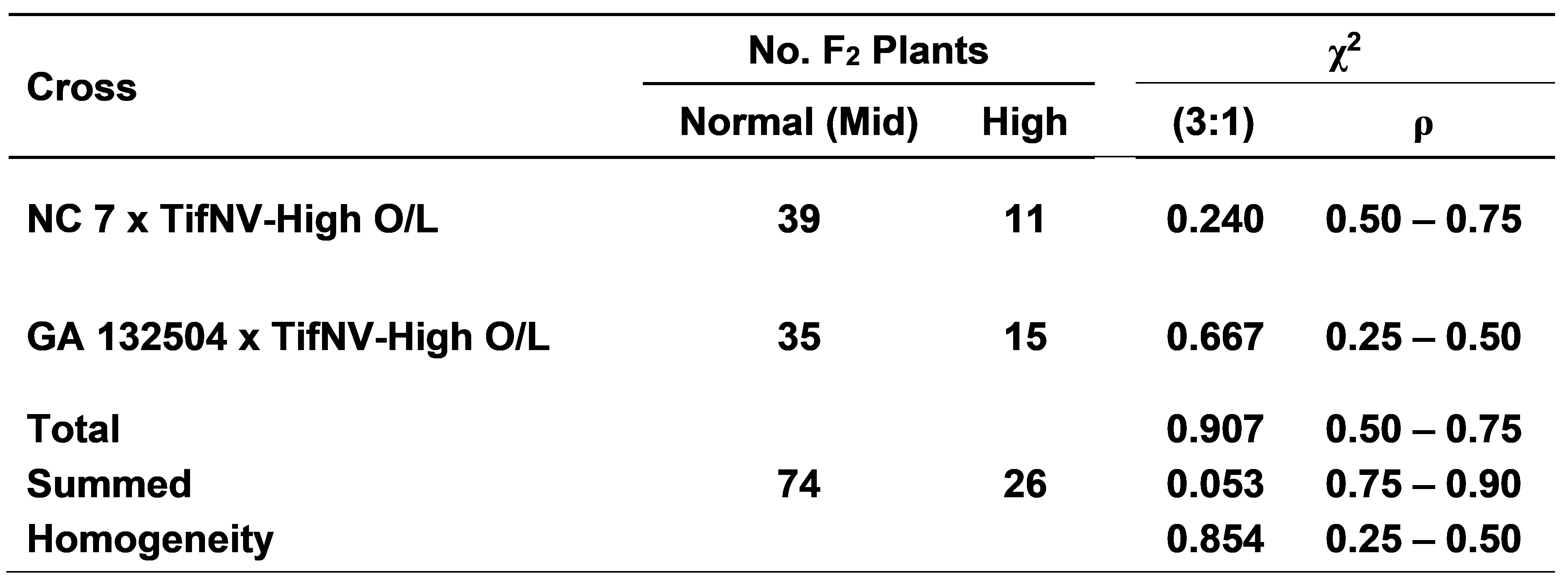
F2 individual plant (F3 seed) segregation for normal (mid) and high-oleic fatty acid ratios from two cross combinations.
The high-oleic trait in peanut has been shown to be controlled by two recessive genes, ol1 and ol2, (Moore and Knauft, 1989; Knauft et al., 1993; and Isleib et al., 1996). Many of the currently available breeding lines and cultivars already carry one of the two recessive alleles for the high-oleic trait. So, it was not unexpected to find a single gene difference with the mid-oleic genotypes used in this study and the high-oleic cultivar, TifNV High-O/L. These inheritance results further support the similarities between mid-oleic and normal-oleic genotypes which are both genetically similar and differ from the high-oleic genotypes (ol1 and ol2) by only one allele. Initially, it was thought that the mid-oleic genotypes might have one of the two recessive genes that was different from the normal-oleic genotypes (ol1 Ol2 x Ol1 ol2). Such cross combinations would have resulted in a 15:1 F2 segregation ratio. Unfortunately, this was not the case.
These results do show that these two mid-oleic genotypes appear to have minor modifier gene(s) that do not involve the high-oleic genes (ol1 or ol2). NC 7 appears to have a completely dominant but weaker minor modifier, while GA 132504 has a stronger but recessive minor modifier. There are many enzymes regulated by different genes involved in peanut seed fatty acid biosynthesis which could explain the differences observed among these two mid-oleic genotypes.
Literature Cited
Ahmed E. M. and Young C. T.. 1982. Composition, quality, and flavor of peanuts. Pp 655-688. In H.E. Pattee and C. T. Young (eds.). Peanut Science and Technology. Amer. Peanut Res. & Educ. Soc., Inc. Yoakum, TX.
Branch W. D. 2007. Registration of ‘Georgia-06G’ peanut. J. Plant Reg. 1: 120.
Branch W. D., Nakayama T., and Chinnan M. S.. 1990. Fatty acid variation among U.S. runner-type peanut cultivars. J. Amer. Oil Chem. Soc. 67: 591-593.
Branch W. D., Brenneman T. B. and Culbreath A. K.. 2003. Tomato spotted wilt virus resistance among high and normal O/L ratio peanut cultivars with and without irrigation. Crop Protection 22: 141-145.
Branch W. D., Perera A. M., and Narayanaswamy K.. 2020. Allelism test between crosses of high-O/L x high-O/L and very high-O/L x very high-O/L peanut genotypes. Peanut Sci. 47: 135-138.
Chu Y., Wu C. L., Holbrook C. C., Tillman B. L., Person G., and Ozias-Akins P.. 2011. Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut. Plant Genome 4: 110-117.
Council for Agricultural Science and Technology. 1991. Food fats and health. Task Force Rept. 118, CAST, Ames, Iowa.
Gurr M. I. and James A. T.. 1975Lipid Biochemistry: An Introduction (2nd Ed.) A Halstead. Press Book, Chapman and Hall, London, and John Wiley & Sons, Inc., New York.
Holbrook C. C., Timper P., Culbreath A. K., and Kvien C. K.. 2008. Registration of ‘Tifguard’ peanut. J. Plant Reg. 2: 92-94.
Holbrook C. C., Ozias-Akins P., Chu Y., Culbreath A. K., Kvien C. K., and Brenneman T. B.. 2017. Registration of ‘TifNV-High O/L’ peanut. J. Plant Reg. 11: 228-230.
Isleib T. G., Young C. T., and Knauft D. A.. 1996. Fatty acid genotypes of five virginia-type peanut cultivars. Crop Sci. 36: 556-558.
Knauft D. A., Moore K. M., and Gorbet D. W. 1993. Further studies on the inheritance of fatty acid composition in peanut. Peanut Sci. 20: 74-76.
Marklund M., Wu J. H. Y., Imamura F., Gobbo L. C. D., Fretts A., deGoede J., P. Shi …and U. Riserus. 2019. Biomarkers of dietary omega-6 fatty acids and incident cardiovascular disease and mortality - An individual level pooled analysis of 30 cohort studies. Circulation 139: 2422-2436.
Moore K. M. and Knauft D. A.. 1989. The inheritance of high-oleic acid in peanut. J. Hered. 88: 252-253.
Mozingo R. W., O’Keefe S. F., Sanders T. H., Hendrix K. W.. 2004. Improving shelf-life of roasted and salted inshell peanuts using high oleic fatty acid chemistry. Peanut Sci. 31: 40-45.
Strickberger M. W. 1968. Chap. 21 Chromosome variation in number. Pp. 449-478. In Genetics. The Macmillian Co., New York, NY.
Wynne J. C., Mozingo R. W., and Emery D. A.. 1979. Registration of NC 7 peanut. (Reg. No. 22) Crop Sci. 19:563.
Notes
-
Author Affiliations
- Professor, Dept. of Crop & Soil Sci., University of Georgia, Tifton, GA 31793 [^]
- Assistant Research Scientist, Dept of Crop and Soil Sci., University of Georgia, Tifton, GA 31793 [^]
- Research Manager, W. M. Keck Metabolomic Research Laboratory, Office of Biotechnology, Iowa State University, Ames, IA 50011-1010 [^] Corresponding author email: wdbranch@uga.edu