

Introduction
It is not possible for every planted peanut ( Arachis hypogaea L.) field to reach an acceptable plant population on the first attempt. Many factors can reduce a plant stand including planting poor quality seed, operating planting equipment improperly (plugged tubes, excessive tractor speed, incorrect depth setting, etc.), planting seed into an environment not suitable for maximized germination (improper soil pH, moisture, or temperature, inadequate seed-soil contact; etc.), failure of a seed treatment to protect against soilborne pathogens, heavy rainfall before emergence, cold water or air temperature fluctuations after planting that rapidly shock the seed, rooting by hogs, or a number of other factors. When an undesirable plant stand occurs, farmers must decide whether to proceed with managing the established crop, abandon the field, destroy the stand and start over to improve correctable soil conditions, or replant the field in an attempt to bolster the plant stand to a suitable population.
There is limited research reported on replant methodology for row crops, especially involving supplemental replanting into a partial plant stand. Previous research has demonstrated the potential for yield improvement by replanting corn ( Zea mays L.) at various population levels with addition of seed to an adjacent furrow parallel to the existing original planted row ( Terry et al., 2012). There is also available information on replanting peanut through supplementation of various initial peanut populations ( Sarver et al., 2016; Sarver et al., 2017). However, when considering this replant management strategy, there are plants at two different developmental stages growing simultaneously in the field. Since peanut is an indeterminate plant, determining the optimum timing for termination is difficult even when all plants are at a uniform growth stage. With plants at two different growth stages, determining the optimum time for digging to maximize yield and grade becomes even more complicated to maximize production so that the earliest planted peanuts don’t become overmature while trying to advance the maturity of the later planted peanuts. Inversely, initiating termination too early could render the replanted plants too immature. In addition, different plant populations could change this dynamic since a drastically poor initial plant stand should result in a larger percentage of replanted plants in order to optimize the final plant stand. A more moderate original plant population may only require a marginal supplemental plant stand to achieve the target population. Therefore, the primary objectives of this research were to quantify yield, grade, and individual plant production characteristics of peanut at multiple sub-optimal plant densities and the effect of supplemental replanting on these variables. An additional objective was to evaluate the timing of termination for the replanted peanut scenarios that correspond with the maturity of the two different plantings.
Materials and Methods
The research was conducted on the University of Georgia (UGA) Lang-Rigdon Farm (31.52, -83.55) in Tifton, GA in the summers of 2014, 2016, and 2017. The experiment was comprised of two primary treatment variables including plant population and replant/termination timing. Plant population included the UGA Extension recommended plant density of 13.1 plants/m as a check treatment ( Baldwin, 1997; Tubbs, 2020). The other plant densities included 3.3, 6.6, or 9.9 plants/m which were each included four times for every replication to correspond factorially with the replant/termination timing treatments. These treatments included 1) no replant, 2) replanted and terminated based on the predicted peanut maturity profile of initial planted peanuts, 3) replanted and terminated based on the predicted peanut maturity profile of the average between the initial and replanted peanuts, and 4) replanted and terminated based on the predicted peanut maturity profile of the replanted plants. This made up a 3 x 4 factorial arrangement plus one positive control, for 13 treatments per replication. A randomized complete block design was used with four replications and four rows in each plot spaced 0.91 m apart with lengths of 9.75 to 10.67 m depending on the size of the field each year. Since two-row equipment was used for planting, management, and harvest activities, the four row plots consisted of two full beds in each plot, one of which was maintained pristine for non-destructive measurements, and the other was used for destructive sampling.
To prepare the field, conventional tillage practices were used by deep turning the soil with a moldboard plow to a depth of approximately 30-35 cm, disk-harrowing, then forming a smooth seedbed with a rotary tiller. Cultivar Georgia-06G ( Branch, 2007) was planted at seeding rates of 22.5 seed/m (for 13.1 and 9.9 plant/m treatments), 15.7 seed/m (for 6.6 plant/m treatments), and 11.1 seed/m (for 3.3 plant/m treatments) on the initial planting date using a single row Monosem NG Plus precision vacuum planter with a PTO driven fan (Monosem Inc., Edwardsville, KS) attached to a McCormick CX105 tractor (McCormick Tractors International Ltd., ARGO SpA, Fabbrico, Italy). After emergence (approximately 3 weeks after planting), plant populations were counted and thinned by hand to specific populations with relatively uniform spacing based on the treatment associated with each plot. Different seeding rates were used at initial planting to ensure an adequate plant stand for the desired population while also reducing the amount of wasted seed and thinning needed compared to using the maximum seeding rate for all plots.
The replant treatments were applied by returning the same tractor/planter to the field approximately 3 weeks after initial planting and offsetting the planter units by approximately 8 cm to the side of the initial row (supplementing the initial plant stand). The unit was always placed to the same side of the initial row to be able to distinguish initial plants from replanted plants for sampling purposes. Since initial plant densities varied by treatment, the replanted seeding rate was adjusted in the attempt to achieve a similar total plant population for all treatments (except for the non-replanted densities, which were used as a set of checks). Original plant populations of 3.3 plants/m were replanted with 15.7 seed/m, populations of 6.6 plants/m were replanted with 12.4 seed/m, and populations of 9.9 plants/m were replanted with 9.1 seed/m. Assumptions had to be made that only a portion of the replanted seeds would survive for each treatment and a larger proportion would survive when replanted next to a lower initial plant density than when competing with a denser initial population. Future research is needed to assess optimum seeding rates for replanted peanuts based on the initial plant density. Maturity was based on the industry standard hull mesocarp color maturity profile method ( Williams and Drexler, 1981). Termination dates were adjusted for inclement weather as needed. All planting, replanting, termination, and harvest dates are included in Table 1 for each year.
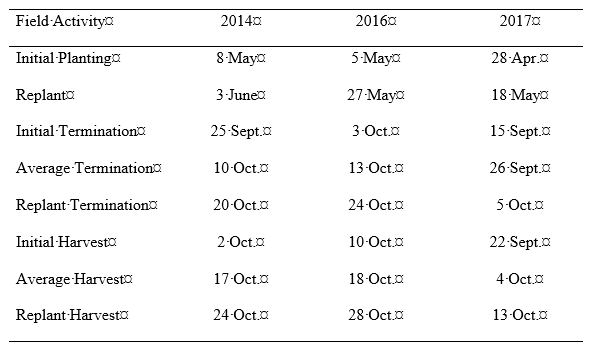
Planting, termination, and harvest dates for each year of plant population/replant/termination timing experiments in Tifton, GA.
Termination was carried out with a KMC digger-shaker-inverter (Kelley Manufacturing Co., Tifton, GA). Roots were severed from the ground with a blade at approximately 15 cm depth and plants were inverted 180 degrees and left in the field for curing until harvest. Harvest was conducted with a two-row KMC peanut combine modified with a side bagging attachment (Kelley Manufacturing Co., Tifton, GA).
Data collected included pod yield, grade (% total sound mature kernels [TSMK]), plant stand, tomato spotted wilt virus (TSWV) incidence, and the number of pods and pod weight per plant for initially planted and replanted peanuts. Yield was adjusted to 7% moisture. Grade was calculated by the Federal State Inspection Service using USDA standards ( USDA-AMS, 2017) as outlined by Davidson et al. (1982). Stand was determined after termination by randomly selecting two segments of 1.5 m length in each of the harvest rows and counting all taproots for initial plants, and replant plants independently. Incidence of TSWV was measured less than 10 days before termination by measuring the total length of row segments in the two harvest rows that had visible symptomology of TSWV infection and converting to a percentage of the total plot length. Incidence of TSWV was not collected in 2017, so was only averaged for 2014 and 2016. The number of pods and pod weights were averaged from five randomly selected plants that were representative of the plot, conducted for initial and replant plants independently.
Statistical analyses were performed by ANOVA using PROC GLIMMIX in SAS 9.4 (SAS Institute Inc., Cary, NC). Year was treated as a random effect. Treatments were pooled over all three years except for TSWV (two years, as noted above), and TSMK partitioned by initial and replanted plants, which was only performed in 2017. Other TSMK analyses were subsamples of the yield, composited of both initial and replanted plants combined. Analyses were conducted in several different ways to gain the best understanding of response depending on the variable being tested. The presence of a stand-alone check made certain analyses unbalanced with it included in the model. To remove bias from supplemental replanting, one analysis was conducted to compare only the four original plant densities (3.3, 6.6, 9.9, and 13.1 plants/m) without influence of replant factors. Some variables (such as comparing replanted plants to initial plants for differences in pod mass/plant and grade) could only be conducted using treatments that included replanting in the model, so those comprised a separate set of analyses. The final analysis was made for comparison of all treatments. This allowed comparison among original plant populations, among termination timings, and also with the positive control. However, this final analysis required every treatment to be considered an independent unit and could not be combined in factorial. The ANOVA results for most of the data are included in Table 2. In each analysis, the LSMEANS statement was used for comparison of treatments (α=0.10). In the combined analyses, mean separations were adjusted using Tukey’s Honestly Significant Different test. Because of the conservative nature of this group comparisons test and the number of treatments being compared, α=0.10 was used as noted by Steel and Torrie (1960). PROC CORR was used to determine correlations among relevant variables. Graphs were prepared with Sigma Plot 14.0 (Systat Software, Inc., San Jose, CA).
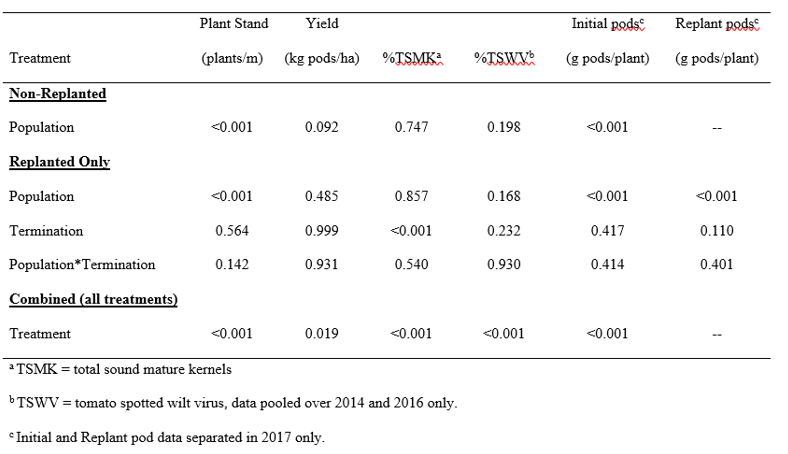
Analysis of variance (level of significance, p values) for four non-replanted plant populations, effect of replanting at three sub-optimal plant populations and three termination timings plus their interaction, and the combined analysis of all treatments as independent units. Data pooled over three years unless noted.
Results and Discussion
Effect of population (without replant)
Comparing the final plant populations of the non-replanted treatments (without interference from replanting), plant stand remained relatively consistent with the original plant population after thinning ( Fig. 1). Plant stands were slightly elevated compared to the original density, which is an artifact of row compression during the digging process. This occurs when the tractor speed is moving slower than the conveyor speed on the inverter. The relative plant stands are most important for treatment comparison, and the slope of a 3.0 plants/m increase in final stand for each 3.3 plants/m increase in original population demonstrates that stands provided the desired separation in plant density throughout the season despite the minor inconsistency in digging speed calibration.
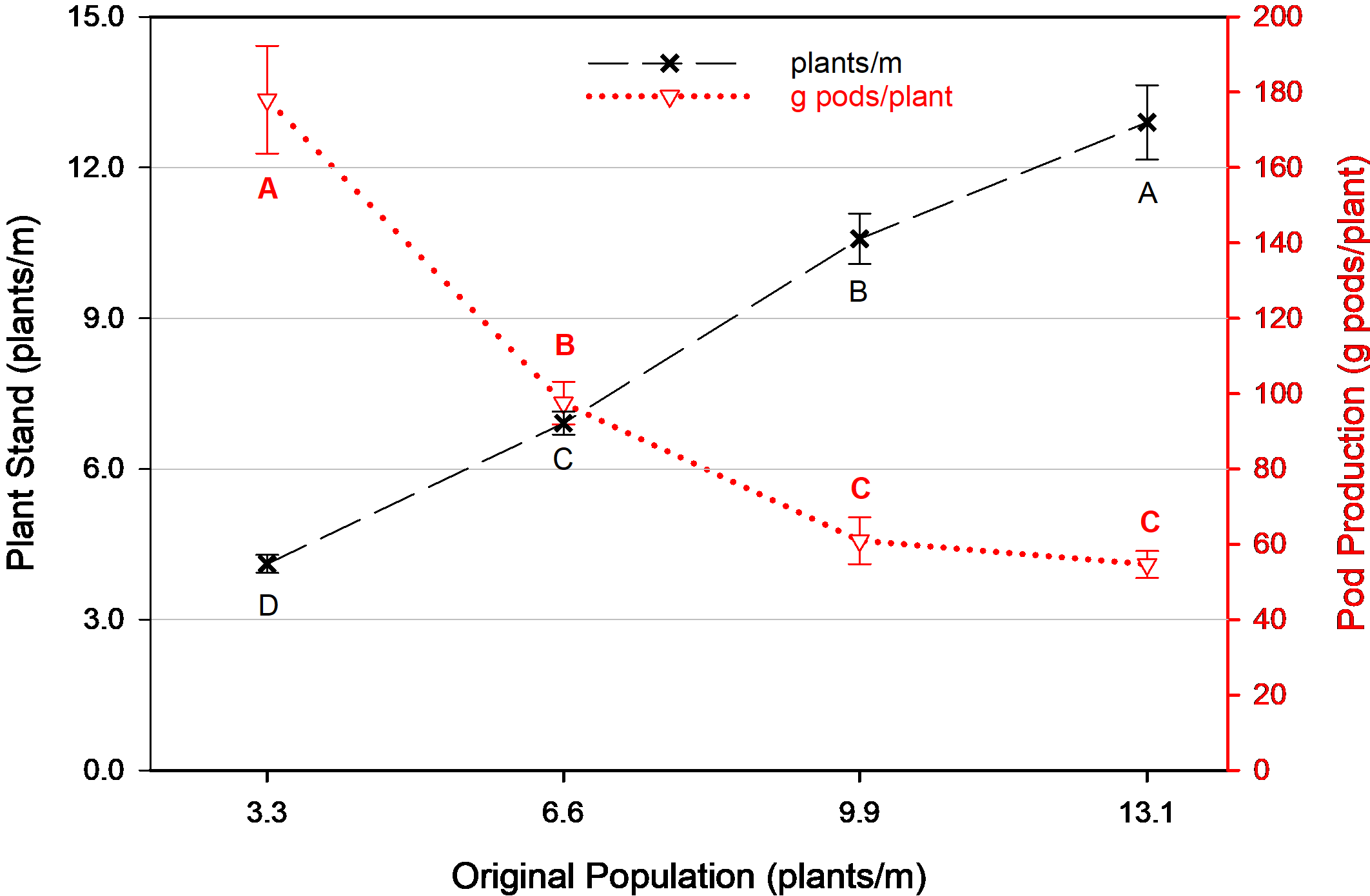
Average plant stand at harvest and individual plant production as influenced by original plant population, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) among populations for each variable. Error bars represent two standard errors of the mean.
Since peanut is an indeterminate plant, it continues to produce pods over the course of the season. When given room to proliferate without competition from neighboring plants (peanut or weeds), the average production on a per plant basis is increased. More space between plants means more pods per plant, which in turn leads to a greater production of pod mass per plant until a plateau ( Sternitzke et al., 2000). A similar response was observed in this research where pod mass per plant was greatest at sparse plant populations, and decreased as plant density increased (from 3.3 to 9.9 plants/m) followed by a plateau (from 9.9 to 13.1 plants/m) ( Fig. 1). Pod mass (g pods/plant) decreased by 45% when stand increased from 3.3 to 6.6 plants/m and was reduced another 38% when stands went from 6.6 to 9.9 plants/m, then plateaued. If evaluated as a linear response where differences were noted (e.g. removing the 13.1 plant/m population since equal to the 9.9 plants/m density), there was a 17 g pods/plant reduction for every 1.0 plant/m increase in population (data not shown).
Yield was significantly different among the plant populations ( Table 2). There was a positive correlation between yield and plant population. The difference between the sparsest and densest populations was 855 kg/ha, which is an 87 kg/ha increase in yield for every additional 1.0 plant/m ( Fig. 2). This is similar to results by Sarver et al. (2016) where a linear increase of 130 kg/ha for each additional 1.0 plant/m was reported for Georgia-06G peanut through the same range of populations.
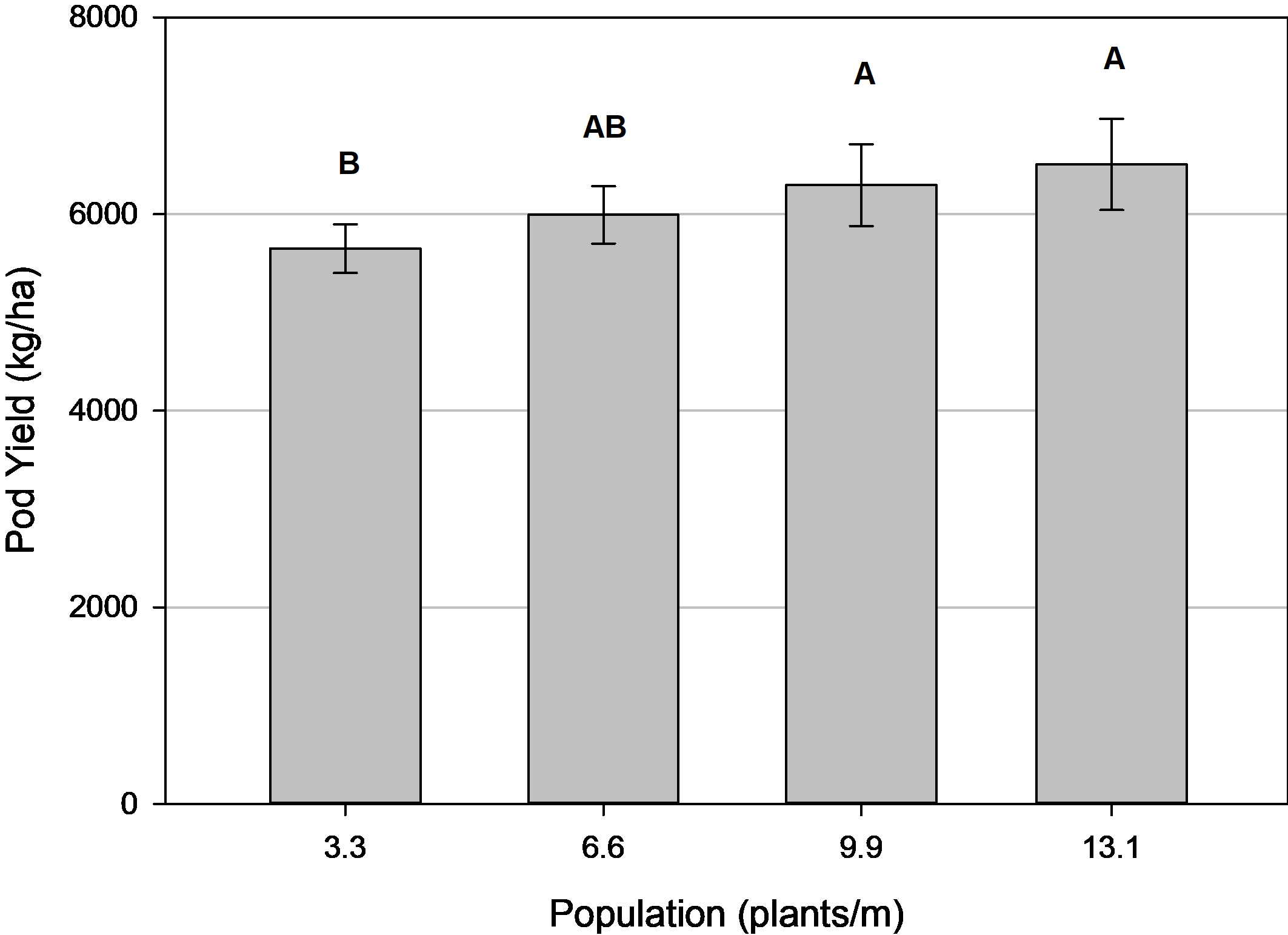
Peanut pod yield as influenced by plant population, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) among populations. Error bars represent two standard errors of the mean.
When there is increased production per plant but fewer plants per row, the advantages/disadvantages from these two situations counteract and cause overall production on an area basis to roughly equilibrate overall. These opposite trends are observed in Fig. 1 and the equilibration is confirmed with a correlation analysis between individual plant production (g pods/plant) and final plant stand (plants/m) that was highly significant (p<0.0001). Hence, overall yield response to plant population was not as pronounced as plant stand since there were no differences in yield between the 3.3 and 6.6 plants/m populations, and similar yield among the 6.6, 9.9, and 13.1 plants/m populations ( Fig. 2). However, it is relevant that yield was greater at the 9.9 and 13.1 plants/m populations compared to the 3.3 plants/m population by 11% and 15%, respectively. This reflects the need for supplemental replanting to increase plant populations and improve yield potential.
Effect of replanting
The second set of analyses removed the non-replanted treatments from the model in order to solely assess the response of populations that were replanted. Similar to the trend observed in the non-replanted peanuts ( Fig. 1), there was a reduction in the pod mass production on an individual plant basis for initial plants ( Fig. 3) even with competition from replanted plants. However, because the replanted plants were also competing for space and other resources, the pod production per plant for each equivalent original population was less in the replanted treatments than in the non-replanted treatments. The reduction in pod mass/plant declined from 17 g pods/plant for each 1.0 plant/m increase with no replant, to merely 7.5 g pods/plant reduction for each 1.0 plant/m increase when accompanied by replanted plants.
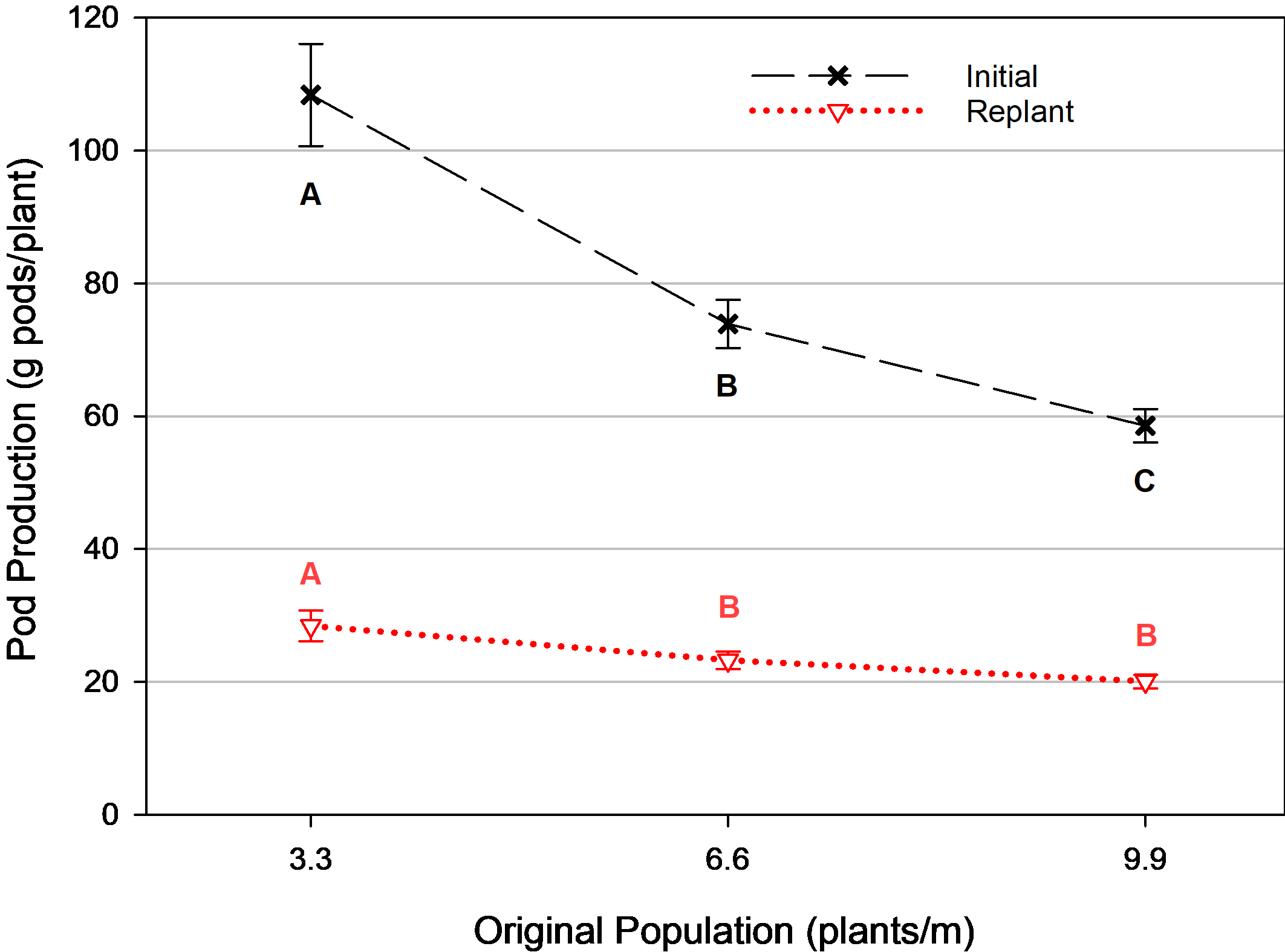
Average pod production for initial and replanted plants by original plant population, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) among populations for each variable. Error bars represent two standard errors of the mean.
A similar, but subtler trend was observed with the replanted plants’ pod production. There was a greater percentage of replanted plants at the smallest original population than at the largest original population overall (data not shown) as a result of the staggered seeding rates in the attempt to make total population more uniform. However, this did not cause the same reduction in pod mass production on a per plant basis from having more plants at a similar growth stage. Since growth of replanted plants was negatively affected from competition by the larger initial plants already in the row, there was increased production for pod mass/plant in the lower original plant populations for replanted plants as well, because of fewer large original plants with which to compete ( Fig. 3). Thus, intra-row plant competition was greater from the original plant population than by other replanted plants. Pod mass of the replanted plants adjacent to the 3.3 plants/m original population was 22% greater than those next to the initial populations of 6.6 plants/m, and 42% greater than those next to the 9.9 plants/m original population.
There were no differences in yield for population or termination timing among the replanted treatments ( Table 2). There was also no difference in grade when the entire composite sample was considered as a whole. However, in the final year of the trial, separate grade samples were collected for initial and replant plants. There were no differences in TSMK from the initial plants at any population. Yet with respect to the replanted plants, grade was greatest in plots where the original plant population was 3.3 plants/m (p=0.041) ( Fig. 4). With less competition from initial plants, the replanted plants in the 3.3 plants/m original population treatment had greater opportunity to grow and develop more rapidly compared to replanted plants that had more competition from initial plants in greater densities.
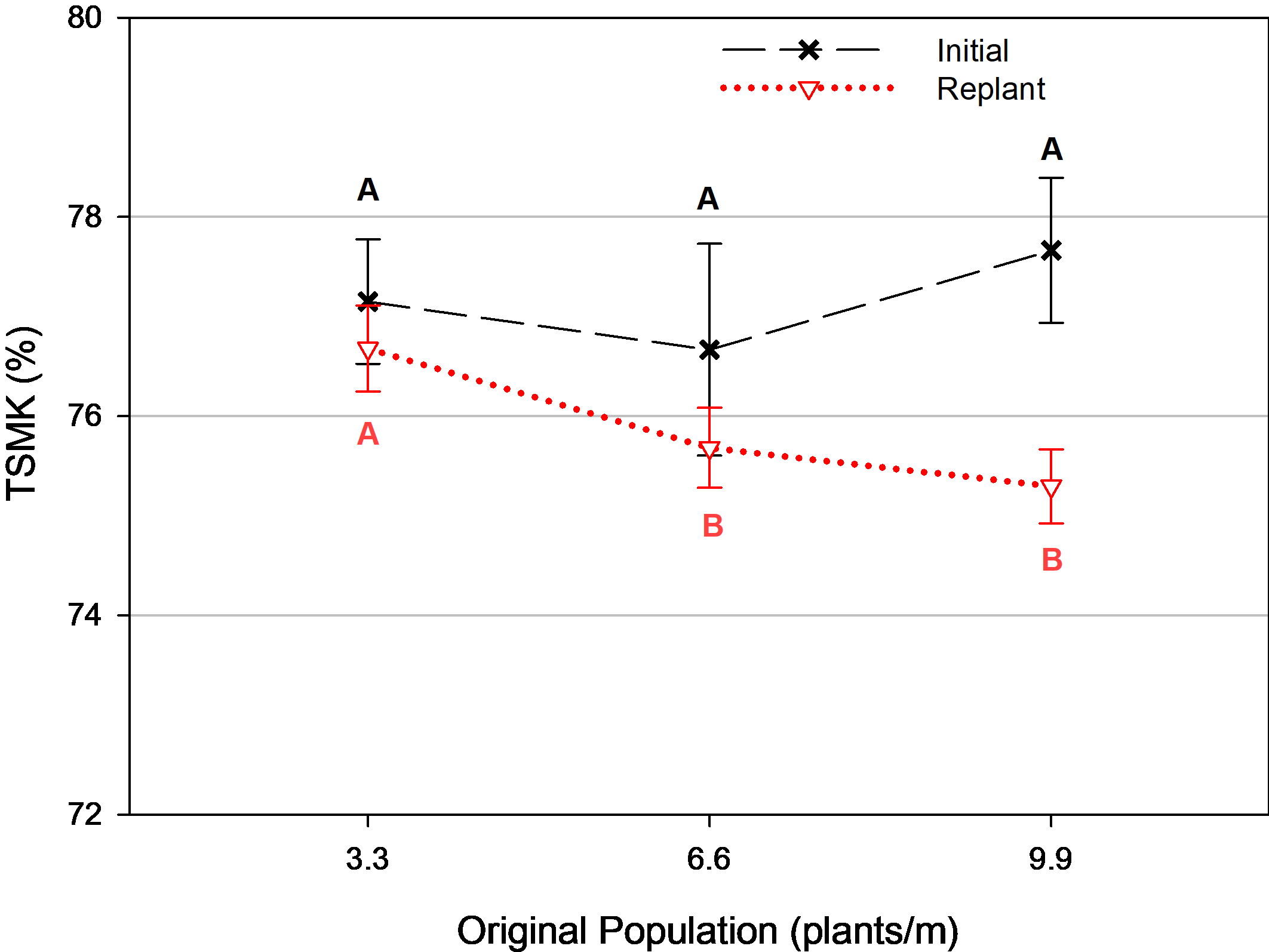
Pod grade (% total sound mature kernels [TSMK]) for initial and replant plants as influenced by original plant population, Tifton, GA, 2017. Means followed by the same letter are not significantly different (α=0.10) among populations for each variable. Error bars represent two standard errors of the mean.
However, there were significant differences in overall grade (initial + replant) based on termination timing ( Table 2). The longer the peanuts were left in the field to mature before termination, the greater TSMK advanced ( Fig. 5). This resulted in approximately 1% increase in TSMK for every additional 5 days the plants were left in the field until the replanted plants reached optimum maturity.
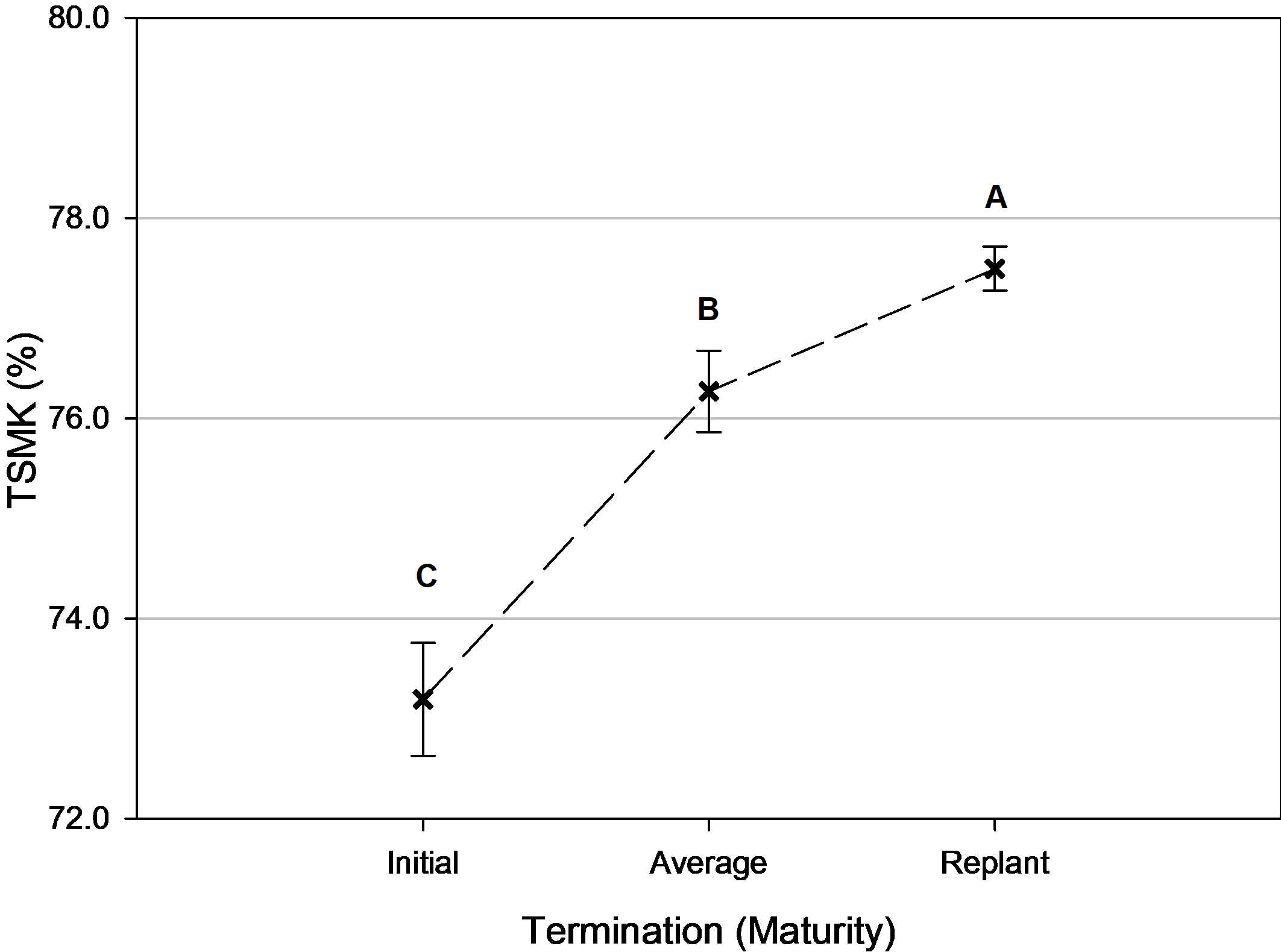
Total pod grade (% total sound mature kernels [TSMK]) as influenced by average termination timing, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) among termination timings. Error bars represent two standard errors of the mean.
When separating the initial and replanted plants for TSMK by termination timing in 2017, the response was different for each. Initial plants were greatest in TSMK at the latest termination date (when replanted plants reached optimum maturity) (p=0.027) ( Fig. 6). However, replanted plants were maximized at the initial termination timing and lowest at the average timing (p=0.072). There is no evidence to explain why this occurred with the replant plants. Further research is needed to investigate this phenomenon using smaller increments of time.

Pod grade (% total sound mature kernels [TSMK]) for initial and replanted plants for termination timings, Tifton, GA, 2017. Means followed by the same letter are not significantly different (α=0.10) among termination timings for each variable. Error bars represent two standard errors of the mean.
Combined analyses of all treatments
At all original plant populations, supplemental replanting next to the original plant stand increased final plant stand ( Fig. 7). This also achieved a combined plant stand equivalent to or greater than the control (13.1 plants/m, Non-replanted) in all replanted treatments. These results show that use of a staggered seeding rate upon supplemental replanting was satisfactory in maintaining the final combined plant stands (initial + replant) at the desired uniformity so that plant population would not be a confounding factor for other variables.
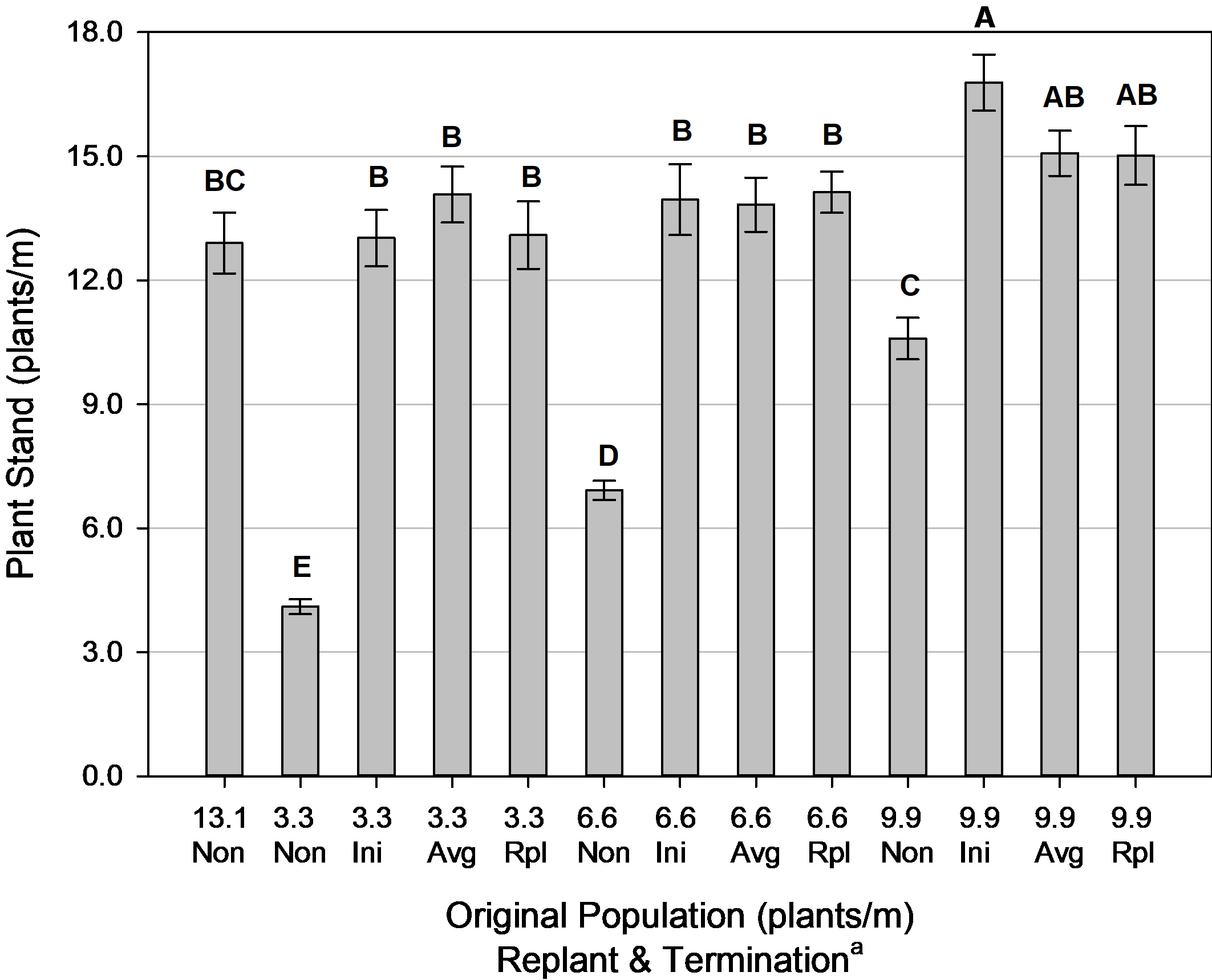
Combined plant stands at harvest (initial + replanted plants) for population and termination, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD test.
a Non = Non-Replanted, Ini = Replanted terminated at Initial Maturity, Avg = Replanted terminated at Average Maturity, Rpl = Replanted terminated at Replanted Maturity.
Yield was positively influenced by replanting at the smallest original plant population and delaying termination until at least the average maturity between initial and replant plants or later ( Fig. 8). However, there was no yield gained from replanting at any other plant population. This is similar to previous research on peanut where yield was improved from supplemental replanting at an initial plant stand of 3.3 plants/m ( Sarver et al., 2016), but not at the other original populations used in this test. Although in that research, there was a yield advantage to supplemental replanting at 8.2 plants/m initial population, yet a yield decline from replanting at 9.8 plants/m.
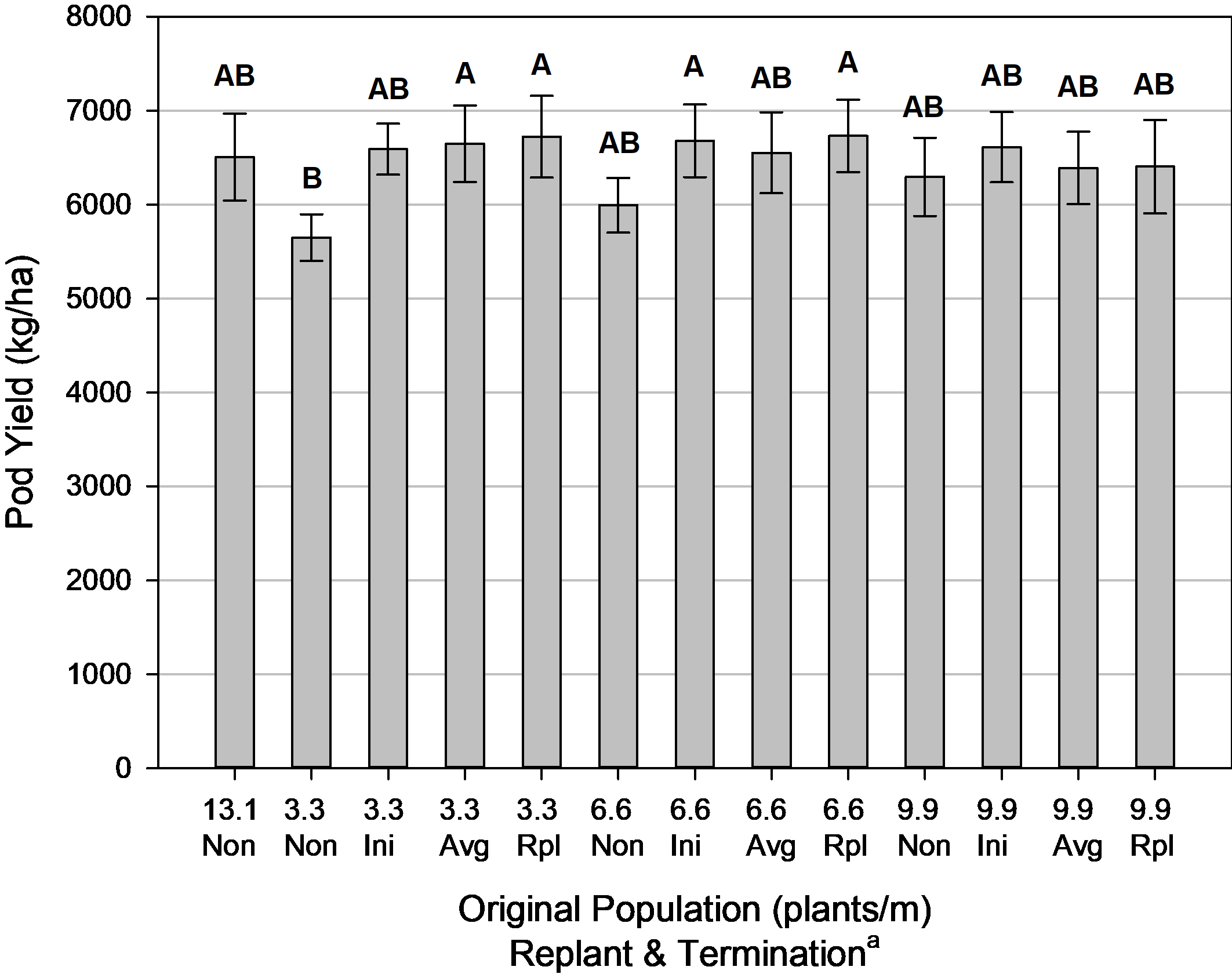
Peanut yield for population and termination, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD test.
a Non = Non-Replanted, Ini = Replanted terminated at Initial Maturity, Avg = Replanted terminated at Average Maturity, Rpl = Replanted terminated at Replanted Maturity.
A similar study on corn also resulted in yield advantages by replanting (at 80,000 seed/ha) when the initial plant populations were extremely low (20,000 plants/ha), and likewise no gain in yield when replanting moderate initial stands (40,000 – 60,000 plants/ha) ( Terry et al., 2012). However, the corn research emphasized that the recommended strategy for that crop would be to destroy the initial stand and start over with a complete replanting of the crop, whereas previous research in peanut suggests that the strategy to destroy and start over will result in yield decline ( Sarver et al., 2016; 2017).
Although, there was a benefit in grade by leaving the plants in the field longer to advance maturity for any given population that was replanted ( Fig. 9). For the 3.3 and 9.9 plants/m original populations, grade was maximized with either the average maturity or replanted plant maturity termination timings. The 6.6 plants/m original population was statistically improved only at the replanted plant maturity, however. Yet, there can be a risk of leaving plants in the ground too long. If the decision to dig is based on the maturity of the replanted plants, then the original plants have pods that are considered overmature. This could lead to pods being left in the ground from the severing of deteriorated pegs or potentially sprouting on the vine if kernels had detached in the hull and dormancy was broken.
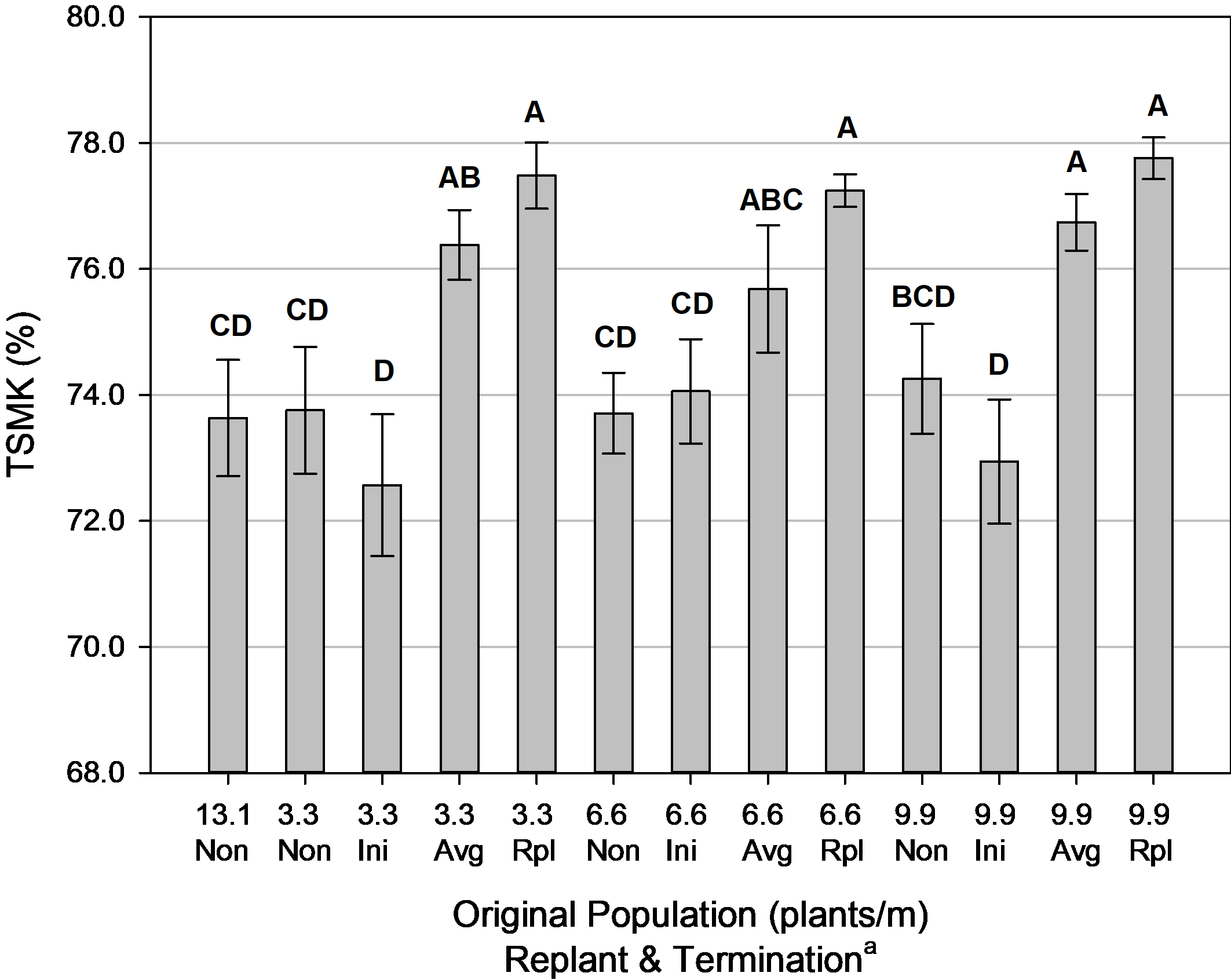
Pod grade (% total sound mature kernels [TSMK]) for population and termination, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD testPod grade (% total sound mature kernels [TSMK]) for population and termination, Tifton, GA, pooled over 3 years (2014, 2016-2017). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD test
a Non = Non-Replanted, Ini = Replanted terminated at Initial Maturity, Avg = Replanted terminated at Average Maturity, Rpl = Replanted terminated at Replanted Maturity.
The average pod mass from initial plants terminated at replant maturity (1.46 g/pod) was less (p=0.005) than those terminated at average maturity (1.56 g/pod) or initial plant maturity (1.55 g/pod). A plausible explanation of this may be related to the largest/densest pods on the original plants were overmature and the pegs broke upon digging, leaving those pods in the ground and reducing the overall average pod mass of the remaining pods. This may not be detected strictly in total grade since the loss of overmature peanuts from initial plants may be offset by continued increasing maturity by the replanted plants. Therefore, while there are benefits to replanting a sub-optimum plant population, careful attention should be made to ensure maximized production at the end of the season so those advantages are not lost from delaying termination too long.
One additional factor that may have contributed to reduced yield in the low population non-replanted treatment was the presence of TSWV ( Fig. 10). There was a strong negative correlation (p=0.0002) between TSWV and yield, displaying a 327 kg/ha reduction in yield for every 10% increase in TSWV. The inclusion of replanted plants helped reduce TSWV incidence in the lower original plant populations (3.3 and 6.6 plants/m) compared to the non-replanted populations. This was not detected in the 9.9 plants/m original plant population though since TSWV was already reduced because of a denser initial plant stand. Reduced incidence of TSWV from the presence of a greater plant stand is similar to observations by Branch et al. (2003), Reddy and Wightman (1988), and Wehtje et al. (1994).
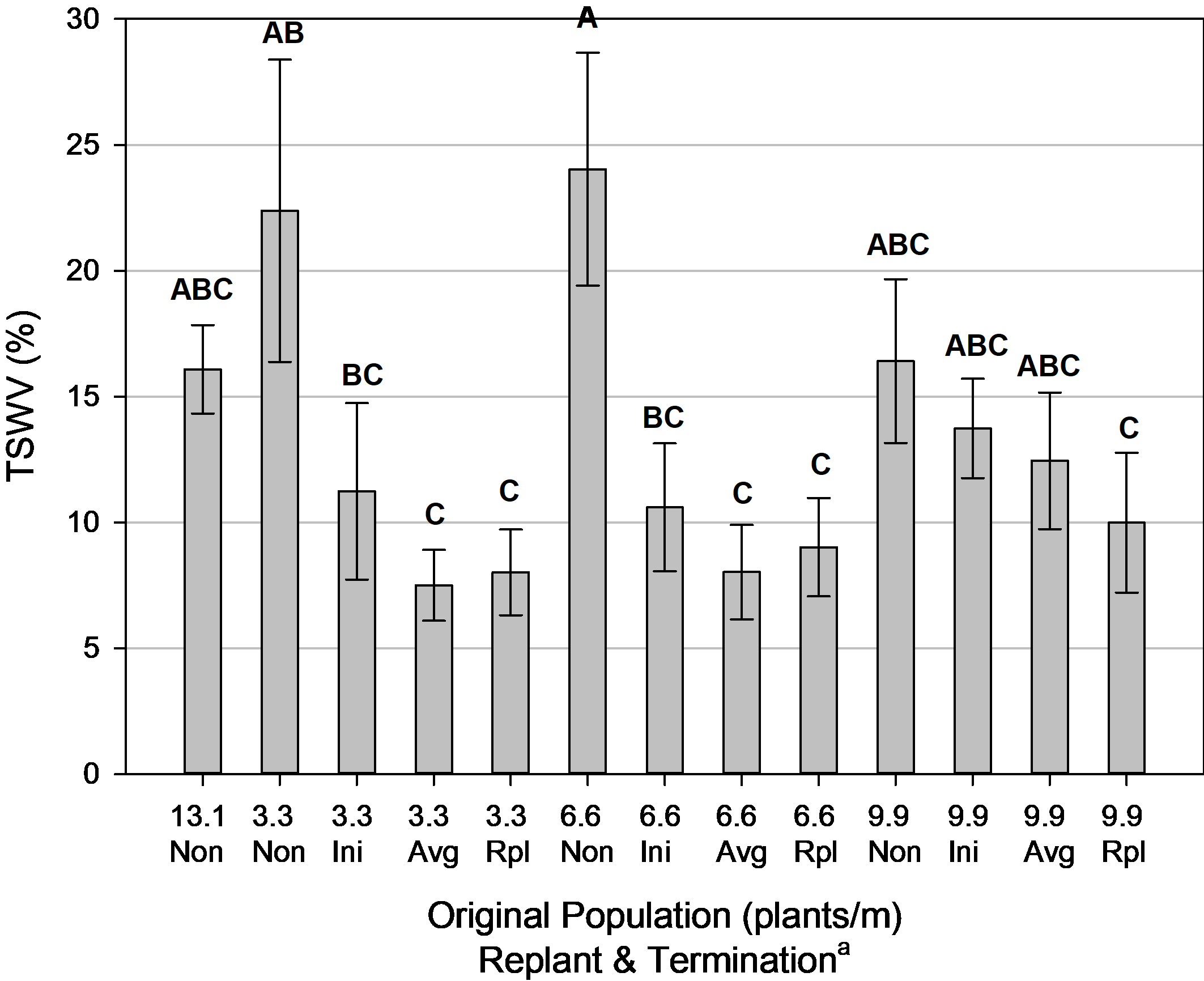
Incidence of tomato spotted wilt virus (TSWV) for population and termination, Tifton, GA, pooled over 2 years (2014, 2016). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD test.Incidence of tomato spotted wilt virus (TSWV) for population and termination, Tifton, GA, pooled over 2 years (2014, 2016). Means followed by the same letter are not significantly different (α=0.10) according to Tukey’s HSD test.
a Non = Non-Replanted, Ini = Replanted terminated at Initial Maturity, Avg = Replanted terminated at Average Maturity, Rpl = Replanted terminated at Replanted Maturity.
Summary and Conclusions
These data suggest that if a farmer has a sub-optimal plant stand after emergence (less than 9.9 plants/m), supplemental replanting next to the original row is a viable option to improve yield and/or grade and reduce the risk of TSWV. There are also advantages to leaving the plants beyond the original plants’ maturity to improve yield and grade up through the replanted plants’ maturity. However, the resulting decreased average pod mass/plant by delaying termination until replant maturity can be risky. Therefore, the recommended digging timeframe for peanuts with supplemental replanting based on these results would be sometime between the average maturity (average of original and replanted plants) and the replanted plants’ maturity for optimum production and minimized risk of losses. This recommendation is based on peanut planting with similar timing to this experiment (original planting within the optimum planting window for peanut). Replanting is not a viable option for increasing yield when the initial planting occurs late in the season (beyond the optimum planting window for peanut) (Sarver and Tubbs, unpublished data). Additional research is needed to optimize timing of replanting and other management decisions such as optimum seeding rate depending on the emerged plant stand. Also, an assessment of variable rate planter technologies and precision agricultural tools to improve the timing and methods of the replant activities is an important component to advancing agricultural practices toward maximum outputs with minimum inputs.
Acknowledgements
The technical efforts of Andy Carter, Nick Liger, and Chris Cromer and the graduate students and student workers who assisted with data collection were essential to the success of this research. We are also grateful to the Georgia Seed Development Commission for providing seed for these experiments, and the National Peanut Board and Georgia Peanut Commission for financial support. This project was conducted in accordance with the objectives of Federal Hatch project #GEO00273.
Literature Cited
Baldwin J.A. 1997. Seeding rates, row patterns and planting dates. pp. 22- 25. In Peanut Production Field Guide. Bull. 1146. Georgia Coop. Ext. Serv. , Athens, GA .
Branch W.D. 2007. Registration of ‘Georgia-06G’ peanut. J. Plant Regis. 1: 120. doi: [: 10.3198/jpr2006.12.0812crc].
Branch W.D., Baldwin J.A., and Culbreath A.K. . 2003. Genotype x seeding rate interaction among TSWV-resistant runner-type peanut cultivars. Peanut Sci. 30: 108- 111.
Davidson J.L., Whitaker T.B., and Dickens J.W. . 1982. Grading, cleaning, storage, shelling, and marketing of peanuts in the United States. pp. 571- 623. In Pattee H.E. and Young C.T. (eds.), Amer. Peanut Res. and Educ. Soc., Inc . Yoakum, TX .
Reddy D.V.R. and Wightman J.A. . 1988. Tomato spotted wilt virus: Thrips transmission and control. pp. 203- 220. In Advances in Disease Vector Research, Vol. 5 . Springer-Verlag, New York, NY 100231 .
Sarver J.M., Tubbs R.S., Beasley Jr. J.P., Culbreath A.K., Grey T.L., Rowland D.L., and Smith N.B. . 2016. Plant population and replant method effects on peanut seeded in single rows. Peanut Sci. 43: 126- 132.
Sarver J.M., Tubbs R.S., Beasley Jr. J.P., Culbreath A.K., Grey T.L., Rowland D.L., and Smith N.B. . 2017. Evaluating plant population and replant method effects on peanut seeded in twin rows. Peanut Sci. 44: 19- 25.
Steel R.G.D., and Torrie J.H. . 1960. Principles and procedures of statistics McGraw-Hill New York, NY. 481 pp.
Sternitzke D.A., Lamb M.C., Davidson J.I., Jr., Barron R.T., and Bennet C.T. . 2000. Impact of plant spacing and population on yield for single-row nonirrigated peanuts (Arachis hypogaea L.). Peanut Sci. 27: 52- 56.
Terry R.M., Dobbels T., Loux M.M., Thomison P.R., and Johnson W.G. . 2012. Corn replant situations: herbicide options and the effect of replanting into partial corn stands. Weed Tech. 26: 432- 437.
Tubbs R.S. 2020. Planting tips. p. 1. In Monfort W.S. (ed.) UGA Peanut Production 2020 Quick Reference Guide. Annual Publ. AP-118. Univ. of Georgia Coop. Ext. , Athens, GA .
USDA-AMS. 2017. Farmers’ stock peanuts inspection instructions. Available at http://www.ams.usda.gov/sites/default/files/media/Farmers%27_Stock_Peanut_Inspection_Instructions%5B1%5D.pdf (verified 10 February 2020). USDA Agric. Marketing Serv., Washington, D.C.
Wehtje G., Weeks R., West M., Wells L., and Pace P. . 1994. Influence of planter type and seeding rate on yield and disease incidence in peanut. Peanut Sci. 21: 16- 19.
Williams E.J., and Drexler J.S. . 1981. A non-destructive method for determining pod maturity. Peanut Sci. 8: 134- 141.
Notes
- Dept. of Crop and Soil Sciences, University of Georgia, Coastal Plain Experiment Station, Tifton, GA 31793. [^] Corresponding author. E-mail tubbs@uga.edu