

Introduction
Peanut ( Arachis hypogaea L.) production worldwide is challenged by yield reduction due to heat stress. Vegetative and reproductive growth and pod yield in peanut are significantly affected by daily average temperatures >30 C ( Wood, 1968; Cox, 1979). Decreases in seed yield by 14, 59, and 90% were reported when day/night temperatures increased from 32/22 C to 36/26, 40/30, and 44/34 C, respectively ( Prasad et al., 2003 ). Daytime temperatures often exceed 30 C during the peanut growing season in the U.S. peanut belt (NCDC, 2021). By the mid-century, air temperature is projected to increase by 1 to 3 C in the U.S., which is expected to cause significant yield declines for all major crops, including peanut (USDA, 2013).
Biological membranes act as the interface between the cell and environment and also compartmentalize metabolism in subcellular organelles. Lipids and proteins are the major constituents of biological membranes. The structure and function of cells are dependent on the fluidity and stability of membranes, which are determined by lipid composition, including the unsaturation levels of fatty acid components ( Zheng et al., 2011 ). Lipids furnish structural, metabolic, and regulatory roles in several aspects of plant growth and development, and responses to environmental stresses ( Welti et al., 2007 ; Okazaki and Saito, 2014; Hou et al., 2016 ). Our previous research identified lipid metabolic changes during heat stress responses in peanut ( Zoong Lwe et al., 2020 ). We profiled the anther lipidome of the cultivars Bailey, Wynne, Georgia-12Y, Sugg, Tifguard, and Phillips, and a germplasm line SPT 06-07 ( Tallury et al., 2014 ), which were exposed to ambient or high temperatures (for 2.5 weeks) during flowering, which is one of the most heat-sensitive stages for peanut. We found that heat stress caused decreases in lipid unsaturation levels in peanut anthers. The decrease in unsaturation level was due to decreases in highly unsaturated lipid species that contained linolenic acid (18:3) and/or increases in less unsaturated lipid species that contained oleic (18:1), linoleic (18:2), or palmitic (16:0) acids. These lipid metabolic changes appeared to be adaptational mechanism in peanut plants to prevent membrane transition into non-bilayer phases and to maintain membrane functionality in anther cells under heat stress.
Peanut seed contains 44 to 56% oil and 22 to 30% protein on a dry weight basis ( Savage and Keenan, 1994). Fatty acid composition of oil is a major seed-quality trait of peanut. Oleic, linoleic, and palmitic fatty acids, together, account for more than 80% of the total fatty acid content in peanut oil ( Dwivedi et al., 1993 ). Storage life and nutritional quality of peanut are largely influenced by the relative proportion of saturated and unsaturated fatty acids in the oil. From a consumer standpoint, increased levels of polyunsaturated and monounsaturated fatty acids in seeds have health benefits ( Reddy et al., 2003 ; Alex et al., 2020 ). However, unsaturation levels are inversely proportional to the storage life of the oil. Specifically, high values for the oleic/ linoleic ratio increase the product stability and shelf-life of peanut oil ( James and Young, 1983; Branch et al., 1990 ). High levels of oleic acid are also beneficial in lowering blood cholesterol ( Reddy et al., 2003). Thus, peanut genotypes with higher levels of oleic acid in seeds have often been used in breeding programs to develop varieties with high oleic to linoleic ratios, which are important for industry ( Branch, 2000; Pattee et al., 2002 ).
Plant heat stress studies have largely overlooked physiological and biochemical alterations in the plant system during the post-stress period ( Jagadish et al., 2020 ). However, very limited information is available on plants’ ability to overcome the damage caused by short episodes of heat stress ( Jagadish et al., 2021 ), which often occurs for a few days to a couple of weeks in the early flowering period of peanut in the U.S. peanut belt (NCDC, 2021). It is not clear whether the heat stress that occurs during an early flowering period will have long-lasting effects on seed composition. If peanut plants employ a memory mechanism after they are exposed to heat stress ( Yamaguchi et al., 2021 ) that includes altering the fatty acid composition in seeds as they do in the reproductive tissues during heat stress (Zoong Lwe et al., 2020), then heat stress will impact seed quality (e.g., high oleic/ linoleic ratio in oil). The objective of the present study was to evaluate the fatty acid composition in seeds collected from peanut genotypes that were exposed to heat stress for a short period (2.5 weeks) at the beginning of the flowering stage.
Materials and Methods
Plant husbandry and heat stress treatment.
The peanut genotypes evaluated in this study were cultivars Bailey, Phillips, and Tifguard and a germplasm line SPT 06-07. In our previous research, Bailey was found to be heat-susceptible, SPT-06-07 to be heat-tolerant, and Tifguard and Phillips to be moderately heat-tolerant based on physiological traits and lipid metabolism (Zoong Lwe et al., 2020). Peanut plants were grown under field conditions at the Clemson University Piedmont Research and Education Center, Pendleton, SC, USA (34°38′51.4″N, 82°43′41.1″W and 260 m above sea.level.) from June to November in 2018 and 2019. The soil type in the experimental site was a Cecil sandy loam (clayey, kaolinitic, thermic typic Hapludults). Field operations followed the recommendations in the Clemson University Peanut Money Maker Guide ( Anco, 2021). Tillage operations included primary tillage using a chisel plow followed by secondary tillage using a rototiller ~8 weeks before planting in 2018 and primary tillage using a rotary plow followed by secondary tillage using a rototiller ~5 weeks before planting in 2019. No fertilizer was applied in either year as the soil was not deficient in any nutrients based on soil tests. Hand-weeding was performed whenever needed in both years. Additionally, pre-emergent herbicides were applied in 2018 to control weeds. Peanut genotypes were sown on 29 June in 2018 and 14 June in 2019 in single-row plots using an Easy-Plant Jab-Type Planter (Product ID: 9178, Johnny's Selected Seeds, Fairfield, ME, USA). Row length was 2.4 m, row spacing was 0.76 m, and plant-plant spacing within each row was 0.05 m in both years. The treatment design was a two-factor factorial based on temperature and genotype. The experiment design was a split plot. The temperature was the whole plot factor with two replications arranged as a randomized complete block design. Genotype was the split-plot factor with four replications arranged as a randomized complete block design within the whole-plot blocks. All plots were maintained as rain-fed throughout the cropping season. No pest or pathogen problems were observed for the duration of the cropping season. The air temperature was monitored every 15 min at the plant canopy level (~ 65 cm from the soil surface) using HOBO data loggers (Onset Computer Corporation, Bourne, MA, USA) from planting through final harvest at maturity (temperature data are reported in Supplementary Figs. S2, S3 of Zoong Lwe et al., 2020).
In both years, heat stress treatment was initiated when the last genotype reached anthesis in both treatments in each block. This was determined by first tagging 10 consecutive plants within each plot and then tagging each of them again when they reached anthesis. Genotypes per treatment per block were considered to have reached anthesis when 50% of the 10 tagged plants in one of the four plots (four replications) reached anthesis ( Boote, 1962). The first and last genotype to reach anthesis differed by ≤7 days in both years. Heat stress treatment was imposed for 17 days in 2018 and 18 days in 2019 using two heat tents. Each heat tent was 4.9 m wide, 13.7 m long, and 3.5 m high. Other details regarding the heat tents and the observed air temperatures inside the heat tents at 65 cm from the soil surface (plant canopy level) are given in Supplementary Figs. S1, S2 and S3 in Zoong Lwe et al. (2020). The average daytime (06:45 to 20:30) temperatures in 2018 during the 17-d treatment period were 41 C inside the heat tents and 31 C outside the heat tents, and the temperatures in 2019 during the 18-d treatment period were 38 C inside the heat tents and 28 C outside. Average night-time temperatures during the treatment period in 2018 were 27 C inside the heat tents and 22 C outside the heat tents, and the night-time temperatures in 2019 were 26 C inside the heat tents and 22 C outside. After the stress treatment period (17 d in 2018 and 18 d in 2019), the greenhouse film covering each heat tent was removed so that all plots remained under ambient temperature until final harvest at maturity. In 2018, a total of 416 mm of rainfall occurred during the growing season and 75 mm during the treatment period. The total rainfall in 2019 was 444 mm during the growing season and 69 mm during the treatment period. The rain did not enter the heat tents while covered with the greenhouse film.
At harvest maturity (growth stage, R8), plants from a 1-m row length were harvested from each plot in 2018. Harvested plants were dried down, and pods were separated using a XH-5000 Dry and Wet Type Peanut Picking Machinery (Henan Xuanhua Imp. & Exp. Trading Co., Ltd., Henan, China). In 2019, plants were harvested from a 1-m row length, dried down, and pods were separated by hand. In both years, ten undamaged healthy pods per plot were randomly selected and seeds were manually separated from pods with care to avoid any physical damage to the seeds. Seeds were air-dried under room temperature and a single undamaged seed per plot was randomly selected for lipid extraction ( Chamberlin et al., 2011 ).
Lipid Extraction from Seeds.
Single seeds per plot were used for lipid extraction following the protocol given by the Kansas Lipidomics Research Center for Arabidopsis ( Arabidopsis thaliana) seeds ( KLRC, 2019) with minor modifications as given below. Each seed was placed in a 50-mL glass tube with a Teflon-lined screwcap (DWK Life Sciences L.L.C., Millville, NJ, USA) that contained preheated, (75 C) 3-mL isopropanol with 0.01% butylated hydroxytoluene. The glass tubes were maintained at 75 C for 15 min to deactivate lipid-hydrolyzing enzymes. After that, each glass tube was cooled to room temperature, and then the seeds were taken out and ground using liquid nitrogen in a mortar & pestle. The ground seed was transferred to a fresh 50-mL glass tube to which 6 mL of chloroform and methanol (1:1), 3 mL of HPLC grade water, and 3 mL of chloroform were added. After that, the tubes were kept in a shaker for 2 hr. The tubes were centrifuged at 1000 rpm for 5 min to obtain separate phases. After transferring the lower layer (chloroform and lipid) to a clean glass tube, 3 mL of chloroform was added to the remaining content. This step was repeated three times, and each time, the clear lower layer was collected in the same glass tube. To the clear solution, 3 mL of 1M KCl solution was added and centrifuged at 1000 rpm for 10 min. The top layer was removed and discarded, and the same step was repeated by replacing 1 M KCl with HPLC grade water. The solvent in lipid extract was evaporated using a N-EVAP 112 nitrogen evaporator (Organomation Associates, Inc., Berlin, MA, USA). Then, the 50-mL glass tube was filled with Nitrogen gas and stored at -20 C. Before shipping the lipid extract to the Kansas Lipidomics Research Center for lipid profiling, the lipid extract in the glass tube was dissolved in 1 mL of chloroform and transferred to a 2-mL clear-glass vial with a Teflon-lined screwcap (DWK Life Sciences LLC., Millville, NJ, USA). The solvent (chloroform) was again evaporated from the 2-mL vials using the nitrogen evaporator and lipid dry weight was determined using a microbalance (AX26, Mettler Toledo, LLC, Columbus, OH, USA) that had a detection limit of 2 μg. Then, the vials containing the lipid extracts were transported to the Kansas Lipidomics Research Center over two days with dry ice.
Fatty Acid Profiling.
At the Kansas Lipidomics Research Center, lipid extracts were used to profile fatty acids following the protocol of Christie (1982). Fifty nanomoles of the internal standard, pentadecanoic acid (15:0) were added to each sample vial containing about 1 mg of lipid extract. After that, 1 mL of 3 M methanolic hydrochloric acid (3 moles of hydrochloric acid in 1 L of methanol solution) was added to each sample, and the samples were bubbled with nitrogen gas and heated at 78 C for 30 min. Afterward, 2 mL of water and 2 mL of hexane were added to each sample. Samples were vortexed and centrifuged to separate the phases, and then the upper hexane phase was removed and saved. This extraction was repeated twice, the solvent was evaporated, and the sample was dissolved in 150 μl of hexane. The samples were then analyzed on an Agilent 6890N gas chromatograph (Agilent Technologies, Wilmington, DE) with a flame ionization detector (FID) which was fitted with a DB-23 capillary column (60 m x 250 μm x 0.25 μm). A sample volume of 1 uL was injected in splitless mode. The gas chromatograph oven temperature was held at 150 C for 1 min and ramped first to 175 C at the rate of 25 C/min and then to 230 C at the rate of 4 C/min, which was held for 9 min. The fatty acids were identified by comparison of retention times of the compounds in the sample with retention times of Supelco 37 component fatty acid methyl ester mix standards [see Table 1 of Sigma-Aldrich/Supelco bulletin 907 ( Sigma-Aldrich, 1996) for a list of the 37 component fatty acids).
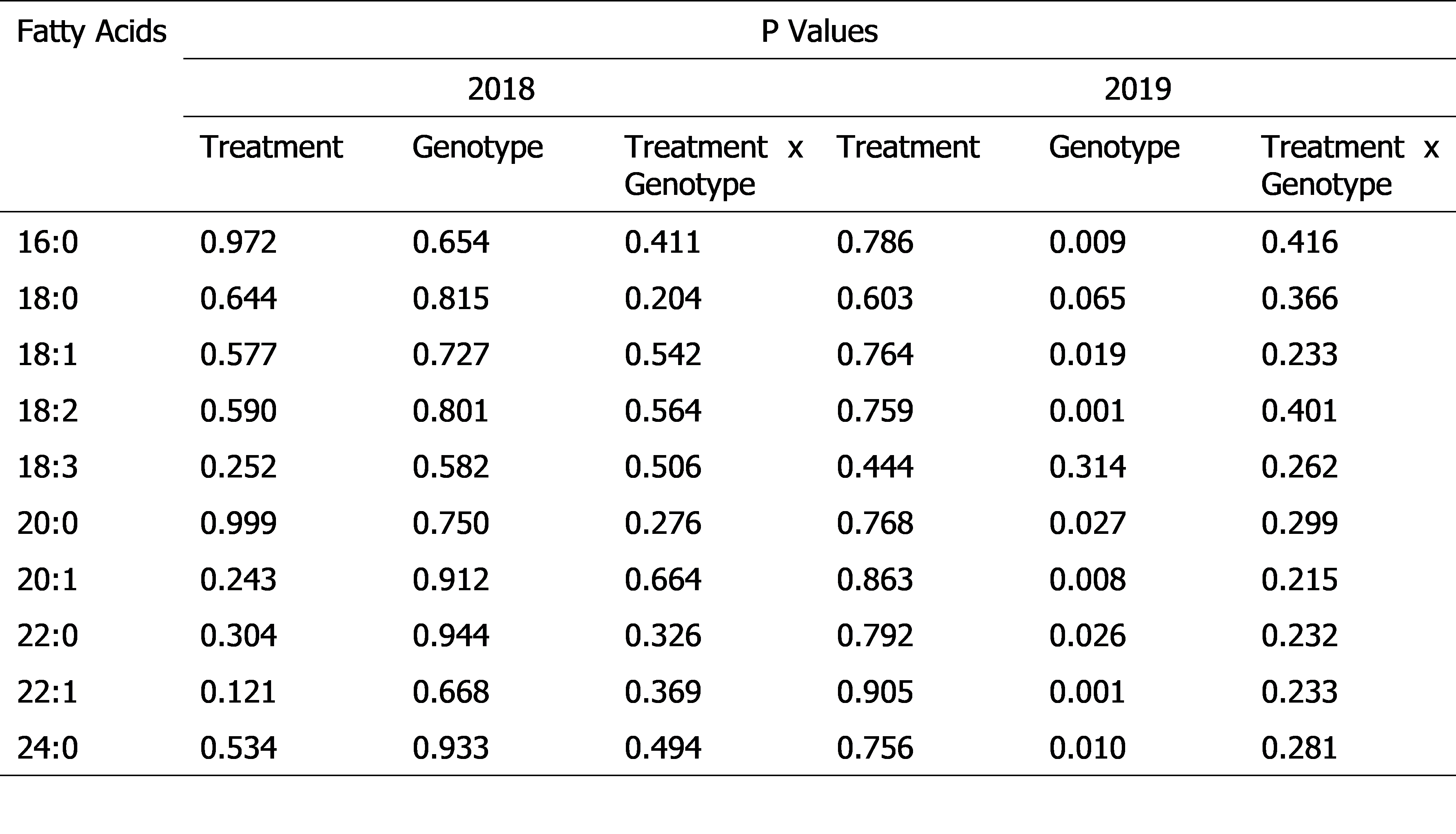
Analysis of variance results on the effects of treatment (ambient temperature and heat stress), genotype ( Bailey, Phillips, and Tifguard in 2018 and Bailey, Phillips, and SPT-06-07 in 2019), and their interaction on fatty acid contents (mg per g of lipid dry weight) of peanut seeds.
Data Analyses.
Analysis of variance and estimation of least-squares means and standard errors were carried out using the GLIMMIX procedure in SAS (Version 9.4, SAS Institute). Treatment and genotype were fixed effects and year and block were random effects. Separation of least-squares means was conducted based on Fisher’s least significant difference (LSD) test (α = 0.05) using the LSMEANS option in the GLIMMIX procedure.
Results and Discussion
The major fatty acids in the seeds of the peanut genotypes were 16:0 (11-12% under ambient temperature and heat stress), 18:1 (43-51% under ambient temperature and 41-49% under heat stress), and 18:2 (30-38% under ambient temperature and 34-40% under heat stress). Other fatty acids were minor and constituted less than 2.7% of the total seed fatty acid content under ambient temperature and less than 2.6% of the total seed fatty acid content under heat stress. The effects of heat stress and heat stress-by-genotype interaction were not significant on seed fatty acid contents in 2018 and 2019 ( Table 1). Peanut genotypes (Bailey, heat-susceptible; SPT-06-07, heat-tolerant; and Phillips, moderately heat-tolerant) did not demonstrate significant alterations in the relative percentage of the major seed fatty acids, 16:0, 18:1, and 18:2 and minor seed fatty acids, 18:0 and 20:0 in both years ( Table 2). Tifguard, another moderately heat-tolerant genotype, demonstrated a decrease in the relative percentage of 18:0 and 18:1 and an increase in the relative percentage of 18:2 and 20:1 in 2018 ( Table 2). However, in 2019, Tifguard did not demonstrate significant alterations in the relative percentage of any seed fatty acid ( Table 2). Future research is warranted to test the anomalous fatty acid alterations of Tifguard in 2018.
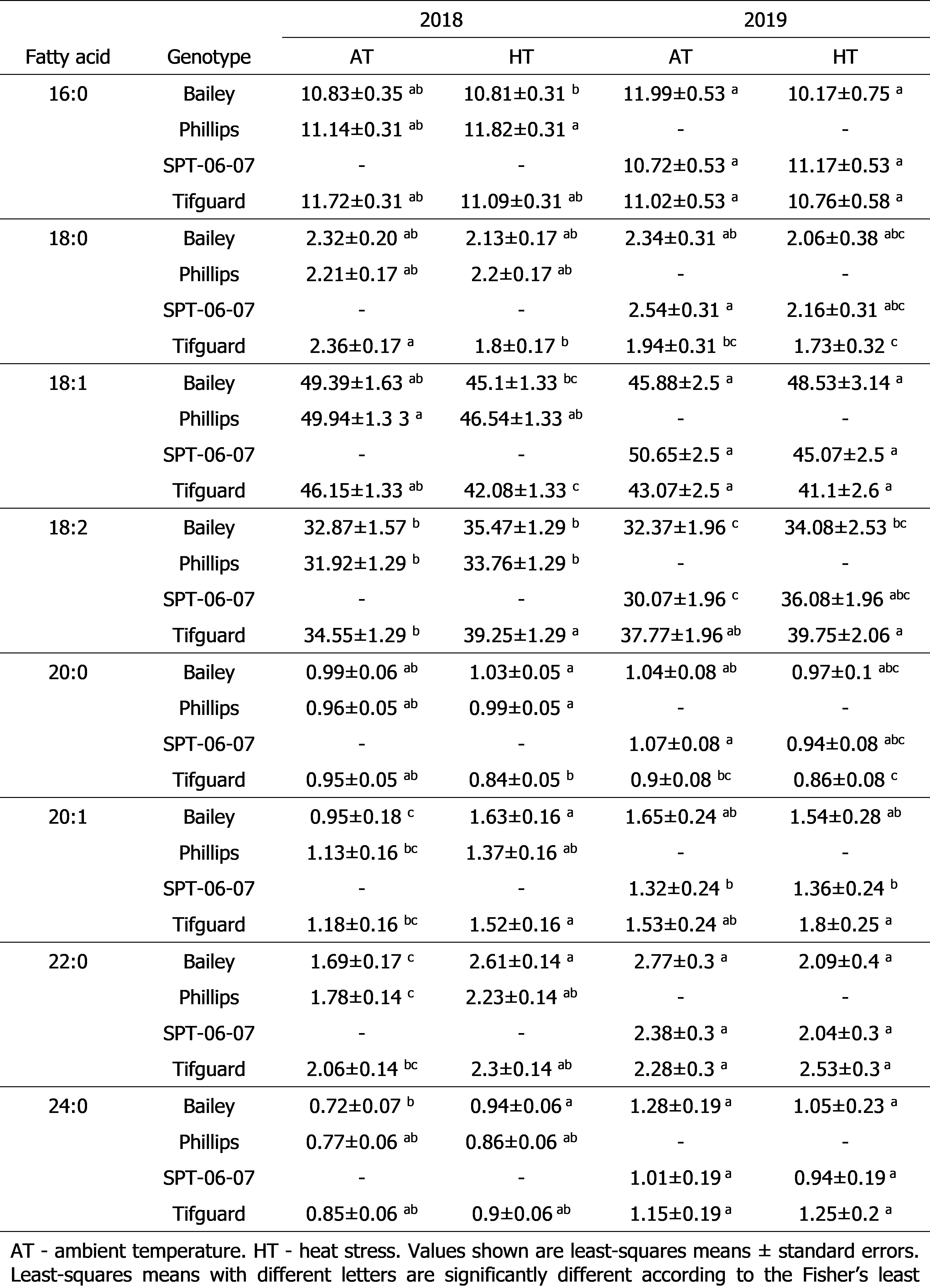
Relative percentage of seed fatty acids under ambient temperature and heat stress for the 2018 and 2019 crop seasons.
In this study, heat treatment was imposed for a short duration (17 d in 2018 and 18 d in 2019) at the beginning of the flowering stage. After that, all plants were maintained under ambient temperature conditions. This means that there was no heat treatment during the seed filling stage. Our results suggest that a short duration of heat stress at the beginning of the flowering stage will not affect oil quality, which is largely influenced by the content of the major seed fatty acids, 18:1 (oleic), 18:2 (linoleic), and 16:0 (palmitic). Instead, oil quality will be more affected if stress occurs during the seed filling/development stage as reported earlier ( Sehgal et al., 2018). It should be noted that the present study did not evaluate any ‘high-oleic’ peanut varieties, and it would be worth confirming the present results on such varieties.
Our results are supported by prior research on oilseed crops ( Dornbos and Mullen, 1992; Reddy et al., 2003 ; Newman et al., 2005 ; Onemli, 2012; Nguyen et al., 2016 ; Sehgal et al., 2018 ). For example, Onemli (2012) and Newman et al. (2005) found that high temperatures during seed development alter seed fatty acid composition and nutritive value of peanut. On the other hand, drought stress early or late in the growing season had little or no effect on seed oil composition in peanut ( Reddy et al., 2003 ). The alteration in the seed fatty acid composition under environmental stresses is believed to be a result of the altered activity of enzymes involved in fatty acid biosynthesis, desaturation, and transport ( Bouchereau et al., 1996 ; Flagella et al., 2002 ; Nguyen et al., 2016 ). For example, oleate desaturase that converts oleic acid (18:1) to linoleic acid (18:2) and linoleate desaturase that converts linoleic acid (18:2) to linolenic acid (18:3) are largely heat-sensitive in most oilseed crops ( Esteban et al., 2004 ; Rolletschek et al., 2007 ; Narayanan et al., 2020 ; Zoong Lwe et al., 2020). The altered activity of oleate desaturase and/or linoleate desaturase can modify the ratio of oleic acid to linolenic acid in oil and thus, affect seed quality. However, based on our results, it appears that peanut plants were able to regain the normal activity of these enzymes by the time of seed filling although they were exposed to heat stress for a short duration at the beginning of flowering.
We recorded pod yield (pod weight m-2) data in 2018 and found that all genotypes demonstrated a decrease in pod yield due to exposure to heat stress ( Fig. 1). Interestingly, in our previous research, we found that genotypes Phillips and Tifguard demonstrated decreases in pod number when they were exposed to heat stress for the same duration (2.5 weeks) at the beginning of the flowering stage (Zoong Lwe et al., 2020. Pod number data were collected from the same experimental plots used for the present study). This may be due to impaired pod-set as heat stress was imposed during flowering when plants set pods (because pod-set occurs when pollen fertilizes the ovule in a flower). Thus, it should be noted that a short duration of heat stress during the early reproductive stage can still affect yield (as it can reduce seed numbers) even though it may not affect seed quality.
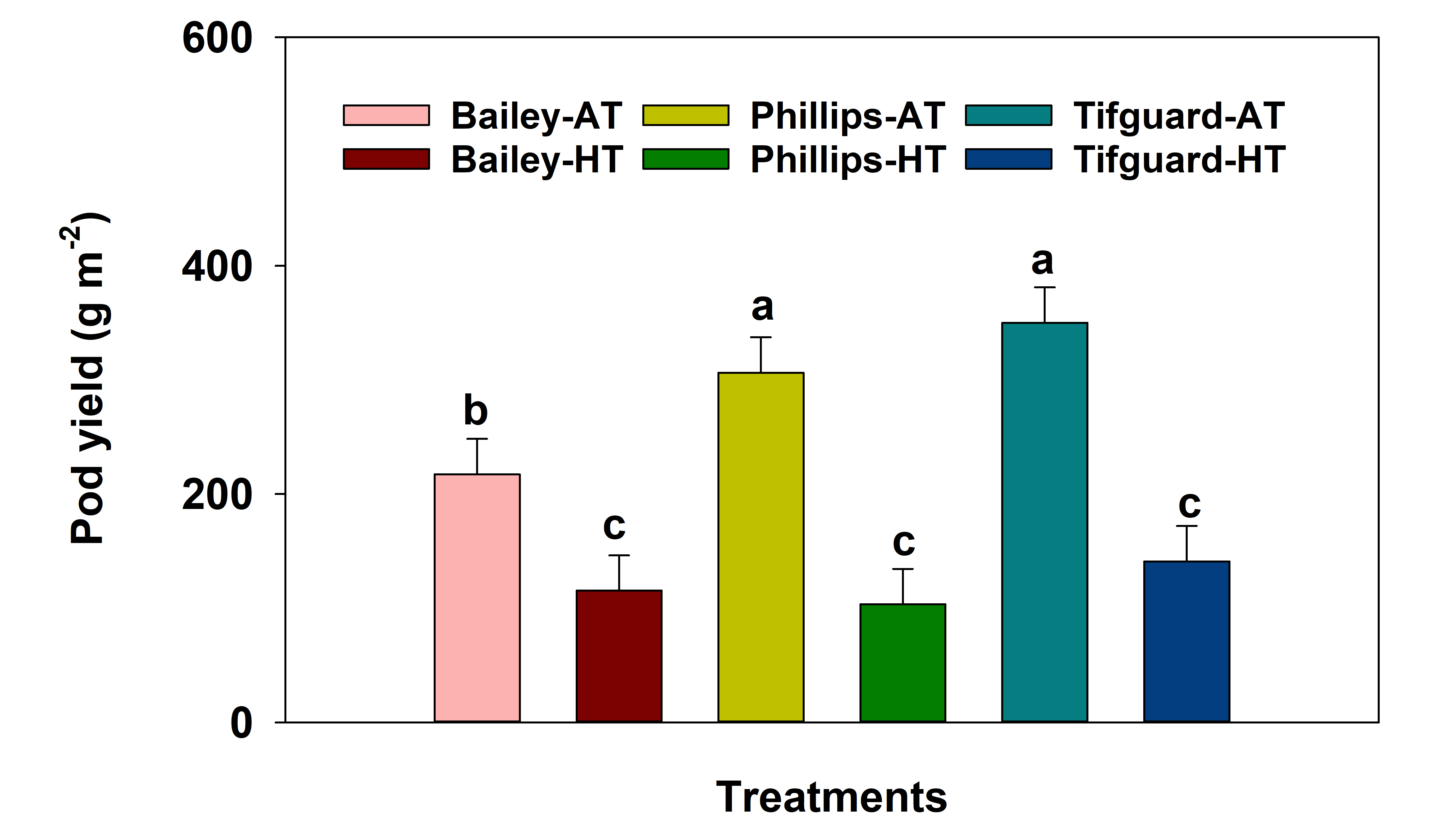
Pod yield of peanut genotypes Bailey, Phillips, and Tifguard under ambient temperature (AT) and heat stress (HT) conditions in 2018. Values shown are least-squares means. Error bars represent standard errors. Least-squares means with different letters are significantly different according to the Fisher’s least significant difference (LSD) test at α = 0.05. Ambient temperature (AT) was 31/22 C (average day/night temperatures during the 17-d treatment period); Heat stress (HT) was 41/27 C. Pod yield was not measured in 2019.
Though the heat stress-by-genotype interaction effect was not significant on seed fatty acid composition in both years, the main effect of genotype was significant on the same parameter in 2019. In 2018, the genotypes sampled were Bailey, Phillips, and Tifguard. While Bailey and Tifguard were also sampled in 2019, Phillips was not evaluated for seed fatty acid content in 2019 due to seed unavailability. Instead, genotype SPT-06-07, which was only grown in 2019 was evaluated for seed fatty acid composition.
The present study only evaluates whether short periods (a couple of weeks) of heat stress that occurred during the beginning of the flowering stage will affect seed fatty acid composition. Since the seeds that we analyzed for the fatty acid composition may or may not have been set and developed during the heat stress treatment, our conclusions are not specific to the effect of heat stress that occurs during the seed development stage.
Single seeds were used in the present study to measure fatty acid composition. As peanuts mature, oleic acid content increases and linoleic acid content decreases in seeds, which suggests that seed maturity affects fatty acid composition ( Pattee et al, 1974; Klevorn et al, 2016; Davis et al., 2017). It should be noted that the variation in seed maturity is not accounted for in the present study. However, the random selection of a seed from each of the four plots for each genotype in each temperature treatment for two years still presents unbiased data for fair comparisons. Furthermore, the dry weight of seeds did not differ among samples (individual seeds representing different treatment-by-genotype-by-replication combinations) (P>0.05) in both years, which might be an indication that the seeds used for fatty acid profiling might be of more or less similar maturity.
Conclusions
The present study evaluated peanut genotypes, Bailey (heat-susceptible), SPT-06-07 (heat-tolerant), Tifguard, and Phillips (moderately heat-tolerant) for the effect of a short-duration (2.5 weeks) heat stress at the beginning of the flowering stage on seed fatty acid composition in a two-year field trial. The results based on the above genotypes showed that a short duration of heat stress at the beginning of the flowering stage did not affect the contents of the major seed fatty acids that affect oil quality (oleic, linoleic, and palmitic acids) regardless of the heat-tolerance level of genotypes. Our results suggest that if heat stress is removed by the time of seed filling, seed fatty acid composition can remain unaffected. However, the heat stress during flowering can decrease pod and seed numbers which will affect yield.
Acknowledgements
We thank the National Peanut Board (Grant # 2014087) for funding this work. This work was supported by the USDA National Institute of Food and Agriculture, Hatch/Multi-State project 1026575. We thank Mr. Ricardo St. Aime, Mr. Enoch Noh, Ms. Jordan Withycombe, Ms. Audrey Jen, Mr. Kevin Wright, Mr. Jerry Zhou, Mr. Andrew McKamy, Mr. Ashwin Koka, Ms. Grace Rhodes, Ms. Pramita Suresh, Ms. Samantha Larson, Ms. Anna Tankersley, Ms. Olivia Mullikan, Ms. Mary Lee, Ms. Elizabeth Middleton, Ms. Kaylex Wilcox, and Mr. Christopher Lebarron for their assistance with field work and data collection. We thank Dr. William Bridges for his consulting time on experimental design and statistical analysis. The fatty acid profiling described in this paper was performed at the Kansas Lipidomics Research Center Analytical Laboratory. Instrument acquisition and method development were supported by the National Science Foundation (including support from the Major Research Instrumentation program; most recent award DBI-1726527), K-IDeA Networks of Biomedical Research Excellence (INBRE) of National Institute of Health (P20GM103418), USDA National Institute of Food and Agriculture (Hatch/Multi-State project 1013013), and Kansas State University. We thank Ms. Libin Yao for conducting gas chromatography with flame ionization detection (FID) to profile fatty acids and Dr. Ruth Welti, Director of the Kansas Lipidomics Research Center, for helps with the same. This paper is Technical Contribution No. 7008 of the Clemson University Experiment Station. The authors declare no conflict of interest.
Literature Cited
Alex A., Abbott K.A., McEvoy M., Schofield P.W., and Garg M.L. . 2020. Long-chain omega-3 polyunsaturated fatty acids and cognitive decline in non-demented adults: a systematic review and meta-analysis. Nutr. Rev. 78: 563- 578.
Anco D. 2021. Peanut money-maker 2021 production guide (ed. Anco, D.). South Carolina State Library.
Bouchereau A., Clossais-Besnard N., Bensaoud A., Leport L., and Renard M. . 1996. Water stress effects on rapeseed quality. Eur. J. Agron. 5: 19– 30.
Branch W.D. 2000. Registration of Georgia Hi-O/L peanut. Crop Sci. 40: 1823.
Branch W.D., Takayama T., and Chinan M.S. . 1990. Fatty acid variation among U.S. runner-type peanut cultivars. J. Am. Oil Chem. Soc. 67: 591- 593.
Boote K.J. 1962. Growth stages of peanut ( Arachis hypogaea L.) . Peanut Sci. 9: 35- 40.
Chamberlin K.D., Melouk H.A., Madden R., Dillwith J.W., Bannore Y., El Rassi Z., and Payton M. . 2011. Determining the Oleic/linoleic Acid Ratio in a Single Peanut Seed: a Comparison of Two Methods. Peanut Sci. 38: 78– 84.
Christie W.W. 1982. Lipid analysis: isolation, separation, identification, and structural analysis of lipids. In: Pergamon international library of science, technology, engineering, and social studies. 2nd ed. Pergamon Press, Oxford. Pp. 52- 53.
Cox F.R. 1979. Effect of temperature treatment on vegetative and fruit growth. Peanut Sci. 6: 14- 17.
Davis J.P., Leek J.M., Sweigart D.S., Dang P., Butts C.L., Sorensen R.B., Chen C.Y., and Lamb M.C. . 2017. Measurements of oleic acid among individual kernels harvested from test plots of purified runner and spanish high oleic seed. Peanut Sci. 44: 134- 142.
Dornbos D.L., and Mullen R.E. . 1992. Soybean seed protein and oil contents and fatty-acid composition adjustments by drought and temperature. J. Am. Oil Chem. Soc. 69: 228- 231.
Dwivedi S.L., Nigam S.N., Jambunathan R., Sahrawat K.L., Nagabhushanam G.V.S., and Raghunath K. . 1993. Effect of genotypes and environments on oil content and oil quality parameters and their association in peanut ( Arachis hypogaea L.) . Peanut Sci. 20: 84- 89.
Esteban A.B., Sicardo M.D., Mancha M., and Martinez-Rivas J.M. . 2004. Growth temperature control of the linoleic acid content in safflower ( Carthamus Tinctorius) seed oil . J. Agric. Food Chem. 52: 332- 336.
Flagella Z., Rotunno T., Tarantino E., Di Caterina R., and De Caro A. . 2002. Changes in seed yield and oil fatty acid composition of high oleic sunflower ( Helianthus Annuus L.) hybrids in relation to the sowing date and the water regime . Eur. J. Agron. 17: 221– 230.
Hou Q., Ufer G., and Bartels D. . 2016. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 39: 1029- 1048.
Jagadish S.V.K., Way A.A., and Sharkey T.D. . 2021. Plant heat stress: Concepts directing future research. Plant Cell Environ. 44: 1992- 2005.
James S.L.H., and Young C.T. . 1983. Comparison of fatty acid content of imported peanuts. J. Am. Oil Chem. Soc. 60: 945- 947.
Klevorn C.M., Hendrix K.W., Sanders T.H., and Dean L.L. . 2016. Differences in development of oleic and linoleic acid in high- and normal oleic virginia and runner-type peanuts. Peanut Sci. 43: 12- 23.
KLRC. 2019. Available at: www.kstate.edu/lipid/analytical_laboratory/protocols_and_methodology/lipid_extraction_arabidopsis_seeds/index.html
Narayanan S., Zoong Z.S. Lwe N. Gandhi R. Welti B. Fallen J.R. , and Rustgi S. . 2020. Comparative lipidomic analysis reveals heat stress responses of two soybean genotypes differing in temperature sensitivity. Plants. 9: 457.
National Climate Data Center (NCDC). 2021. https://www.ncdc.noaa. gov (Accessed 08/13/2021).
Newman Y.C., Sollenberger L.E., Boote K.J., Allen L.H., Jr., Vu J.C.V., and Hall M.B. . 2005. Temperature and carbon dioxide effects on nutritive value of rhizoma peanut herbage. Crop Sci. 45: 316- 321.
Nguyen Q.T., Kisiala A., Andreas P., Emery R.J.N., and Narine S. . 2016. Soybean seed development: fatty acid and phytohormone metabolism and their interactions. Curr. Genomics. 17: 241- 260.
Okazaki Y., and Saito K. . 2014. Roles of lipids as signalling molecules and mitigators during stress response in plants. Plant J. 79: 584- 596.
Onemli F. 2012. Impact of climate change on oil fatty acid composition of peanut ( Arachis hypogaea L.) in three market classes . Chil. J. Agri. Res. 72: 483- 488.
Pattee H.E., Johns E.B., Singleton J.A., and Sanders T.H. . 1974. Composition changes of peanut fruit parts during maturation. Peanut Sci. 1: 57- 62.
Pattee H. E., Isleib T.G., Gorbet D.W., Moore K.M., Lopez Y., Baring M.R., and Simpson C.E. . 2002. Effect of the high-oleic trait on roasted peanut flavor in backcross-derived breeding lines. J. Agri. Food Chem. 50: 7362- 7365.
Prasad P. V. V., Boote K.J., Allen L.H., Jr., and Thomas J.M.G. . 2003. Super-optimal temperatures are detrimental to peanut ( Arachis hypogaea L.) reproductive processes and yield at both ambient and elevated carbon dioxide . Glob. Chang. Biol. 12: 1775- 1787.
Reddy T.Y., Reddy V.R., and Anbumozhi V. . 2003. Physiological responses of groundnut ( Arachis hypogea L.) to drought stress and its amelioration: a critical review . Plant Growth Regul. 41: 75- 88.
Rolletschek H., Borisjuk L., Sanchez-Garcia A., Gotor C., Romero L.C., Martinez-Rivas J.M., and Mancha M. . 2007. Temperature-dependent endogenous oxygen concentration regulates microsomal oleate desaturase in developing sunflower seeds. J. Exp. Bot. 58: 3171- 3181.
Savage G.P., and Keenan J.I. . 1994. The composition and nutritive value of groundnut kernels. In: Smartt J. (eds) The Groundnut Crop. World Crop Series. Springer, Dordrecht. p. 173- 213
Sehgal A., Sita K., Siddique K.H.M., Kumar R., Bhogireddy S., Varshney R.K., Rao B.H., Nair R.M., Prasad P.V.V., and Nayyar H. . 2018. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 9: 1705.
Sigma-Aldrich. 1996. Comparison of 37 component FAME standard on four capillary GC columns. Sigma-Aldrich/Supelco Bulletin 907.
Tallury S.P., Isleib T.G., Copeland S.C., Anderson P.R., Balota M., Singh D., and Stalker H.T. . 2014. Registration of two multiple disease-resistant peanut germplasm lines derived from Arachis cardenasii Krapov. & W.C . Gregory, GKP [: 10017. J. Plant Regist. 8:86-89].
United States Department of Agriculture (USDA). 2013. Climate change and agriculture in the United States: Effects and adaptation. Technical Bulletin p. 1935.
Yamaguchi N., Matsubara S., Yoshimizu K., Seki M., Hamada K., Kamitani M., Kurita Y., Nomura Y., Nagashima K., Inagaki S., Suzuki T., Gan E.S., To T., Kakutani T., Nagano A.J., Satake A., and Ito T. . 2021. H3K27me3 demethylases alter HSP22 and HSP17.6C expression in response to recurring heat in Arabidopsis . Nat. Commun. 12: 1.
Welti R., Shah J., Li W., Li M., Chen J., Burke J.J., Fauconnier M.L., Chapman K., Chye M.L., and Wang X. . 2007. Plant lipidomics: discerning biological function by profiling plant complex lipids using mass spectrometry. Front. Biosci. 12: 2494- 2506.
Wood I.M.W. 1968. The effect of temperature at early flowering on the growth and development of peanut ( Arachis hypogaea L.) . Aust. J. Agr. Res. 19: 241- 251.
Zheng G., Tian B., Zhang F., Tao F., and Li W. . 2011. Plant adaptation to frequent alterations between high and low temperatures: Remodelling of membrane lipids and maintenance of unsaturation levels. Plant Cell Environ. 34: 1431- 1442.
Zoong Lwe Z.S., Welti R., Naveed S., Rustgi S., Anco D., and Narayanan S. . 2020. Heat stress elicits remodelling in the anther lipidome of peanut. Sci. Rep. 10: 22163.