

Introduction
Early leaf spot (ELS) and late leaf spot (LLS) caused by Passalora arachidicola (Hori) U. Braun (syn. Cercospora arachidicola) and Nothopassalora personata (syn. Cercosporidium personatum (Berk. & Curt.) Deighton), respectively, are foliar fungal diseases of peanut (Arachis hypogaea L) responsible for significant yield losses. Wild Arachis species are potential sources of resistance to various peanut diseases including leaf spots (Abdou et al., 1974; Stalker, 1984; Singh, 1986; Stalker and Moss, 1987; Tallury et al., 2014a; Stalker et al., 2017; Arias et al, 2018). Among the 81 wild species described in the genus, only taxa within section Arachis cross readily with cultivated peanut. Therefore, several studies have been focused on the screening of species within section Arachis and a number of resistant sources have been reported (Subrahmanyan et al., 1985; Pande and Rao, 2001; Fávero et al., 2009; Michelotto et al., 2015). Most of these studies have been conducted in greenhouses or laboratory. However, screening of germplasm under natural infestation for a large number of accessions has been limited.
High levels of resistance or immunity to leaf spots have been identified in the wild diploid species Arachis cardenasii Krapov. & W.C. Gregory, A. diogoi Hoehne, and A. stenosperma Krapov. & W.C. Gregory (Stalker et al., 1979; Simpson, 2001; Stalker et al., 2017). These species have been extensively used in interspecific crosses to develop breeding materials for introgression. Multiple introgression lines from a A. hypogaea × A. diogoi interspecific derived population have demonstrated the potential of using wild Arachis species for peanut improvement (Hancock et al., 2019). Arachis cardenasii derived introgression lines have been utilized as a major source of multiple disease resistances, including ELS (Chu et al., 2019; Chu et al., 2020; Stalker et al., 2002; Tallury et al., 2014b). More recently, major quantitative trait loci for resistance to early and late leaf spot diseases have been identified in a recombinant inbred line population (Florida-07 × GP-NC WS16) (Chu et al., 2019). The resistant parental line, GP-NC WS16, is an interspecific breeding material with introgression from A. cardenasii (Stalker and Beute, 1993; Tallury et al., 2014b). In the present study, 78 accessions representing 15 species of section Arachis from the U.S peanut germplasm collection were screened for resistance to leaf spots. The objective was to identify potential candidates for interspecific hybridization and pre-breeding.
Materials and Methods
A total of 78 accessions representing 15 wild species of Arachis section Arachis, including 72 diploids (2n = 2x = 20) and 6 tetraploids (2n = 4x = 40) from the U.S. peanut germplasm collection were evaluated at the USDA-ARS National Peanut Research Laboratory at Dawson, Georgia, during 2017 and 2018. Additionally, six peanut runner cultivars, three susceptible (‘Georgia-13M’, ‘TUFRunner 511’, ‘Georgia-09B’) and three resistant cultivars (‘Georgia-14N’, ‘Georgia-06G’, ‘TifNVHigh OL’) were used as checks (Table S1). Seeds were planted in the greenhouse in 5.7 cm × 5.7 cm peat pots containing peat-based potting mix. After 5-6 weeks, plants were transplanted into the field at the Newman and Bolton farms in 2017 and 2018 growing seasons, respectively. The experimental design was a randomized complete block with two replications. Each block consisted of a single row with five plants separated 0.61 m apart and 1.83 m between rows. Plants were grown using common agricultural practices for peanut production in Southwest Georgia with supplemental irrigation. No fungicides were applied for prevention or control of leaf spots. Chemicals for weed and insect control were applied following manufacturer’s recommendations. Weeds were also controlled by manual hoeing. Meteorological data were collected at the Newman farm, Terrell County, Georgia. This weather station was used as a reference for both farms (www.weather.uga.edu).
Screening was conducted under field (natural inoculum) conditions. Disease severity was evaluated by final defoliation rating (FD), early leaf spot lesion counts (ELC), and late leaf spot lesion counts (LLC). Defoliation was rated on whole plots at the end of the season using the proportional 1-9 scale, with 1 indicating no defoliation and 9 complete defoliation (Chiteka et al., 1988; Tallury et al., 2009). For lesion counts, five leaves were randomly chosen from a lateral branch of each plant in the plot. Lesions were examined under a stereomicroscope and the number of lesions were counted for ELS and LLS, separately. Images of leaf spot lesions were captured with a stereomicroscope Leica MZ16F (Leica, Wetzlar, Germany).
Data from FD, ELC, and LLC were subjected to analysis of variance (ANOVA), followed by Tukey’s test for mean comparisons. Variables were transformed to the lambda power of 0.5 (FD) and log10 (ELC, LLC) to meet the assumptions of ANOVA. Phenotypic correlations among variables were calculated and plotted using the corrplot package (Wei et al., 2017). All statistical analyses were conducted in R environment (R Development Core Team, 2020).
Results and Discussion
Environmental conditions in 2017 and 2018 growing seasons (May-October) were favorable for leaf spot development, as indicated by the severe leaf spot symptoms observed in both susceptible and resistant checks (Table 1). Average maximum and minimum temperatures showed a similar trend both years, with the exception of June (maximum) and September (minimum/maximum) 2018, which were significantly higher (P < 0.01) (Fig. 1).
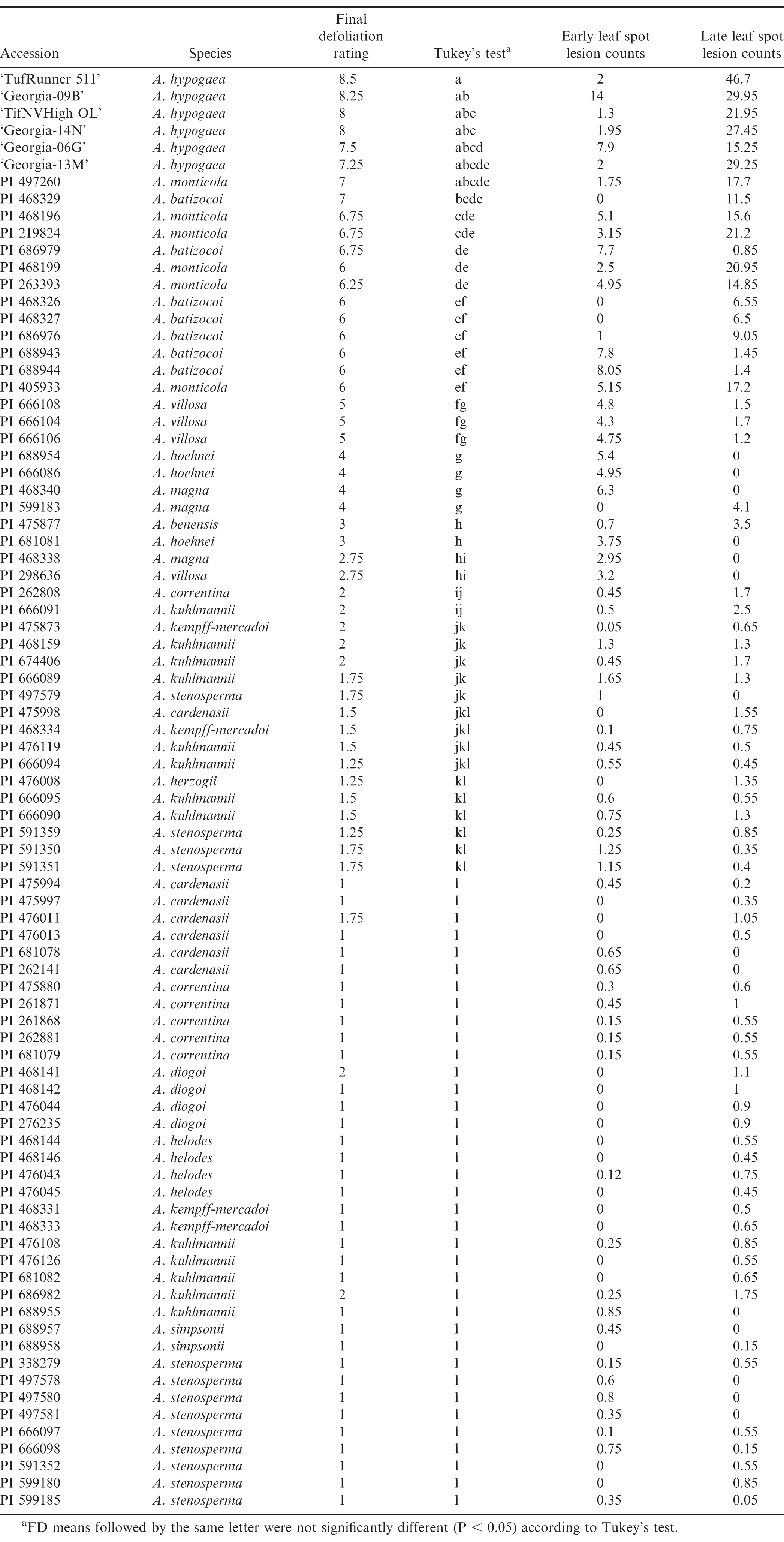
Accession means for all disease variables measured on 78 wild Arachis accessions and five runner cultivar checks.
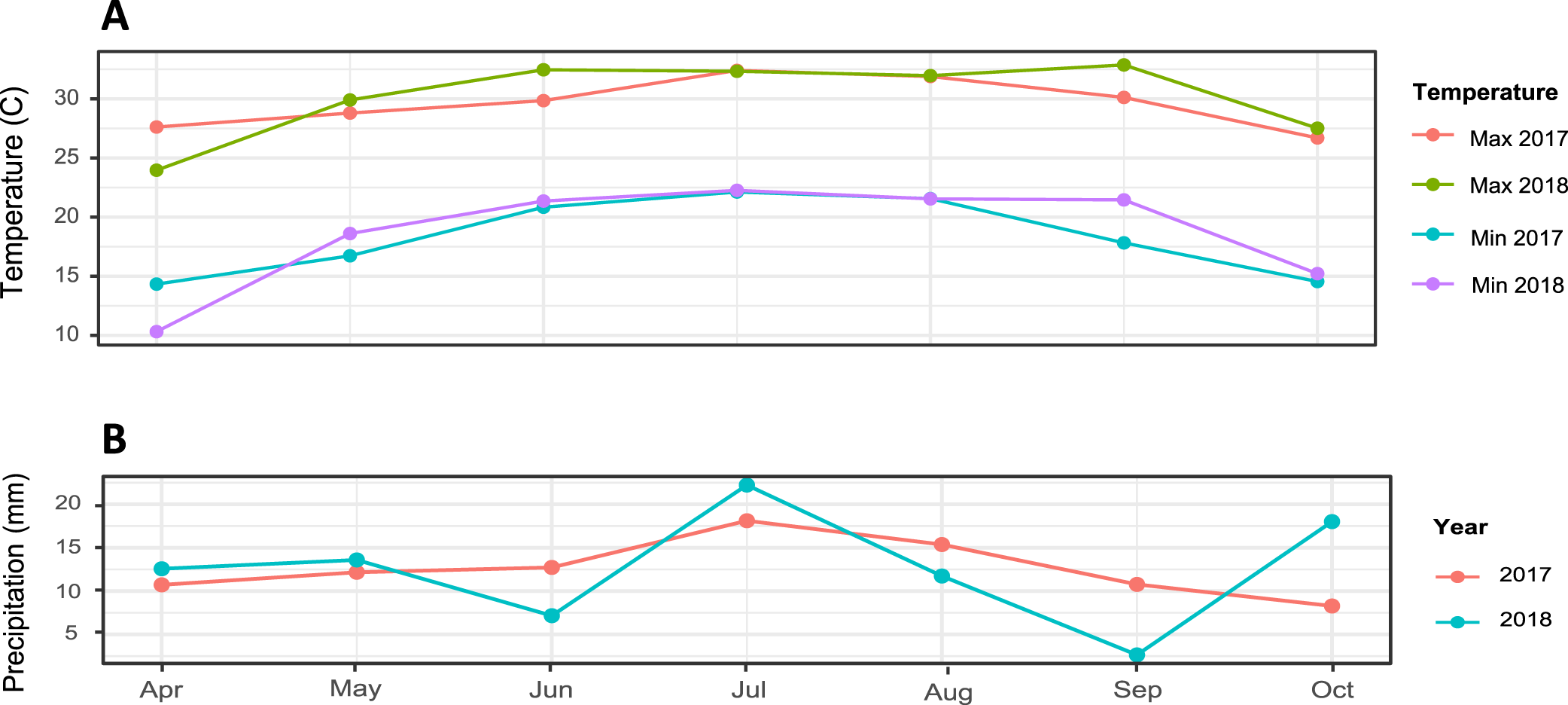
(A) Average maximum and minimum temperatures and (B) cumulative monthly precipitation at Newman, Terrell County, Georgia weather station.
Correlation coefficients between variables were estimated based on the mean of two years. Final defoliation ratings were highly correlated (r = 0.92) with the total number of lesions (ELC + LLC) and moderately correlated (r = 0.64-0.66) with ELC and LLC when analyzed independently (Fig. 2). Results further showed that LLS was predominant in 70% of accessions, which can explain the low correlation (r = 0.14) between ELC and LLC (Fig. 2).
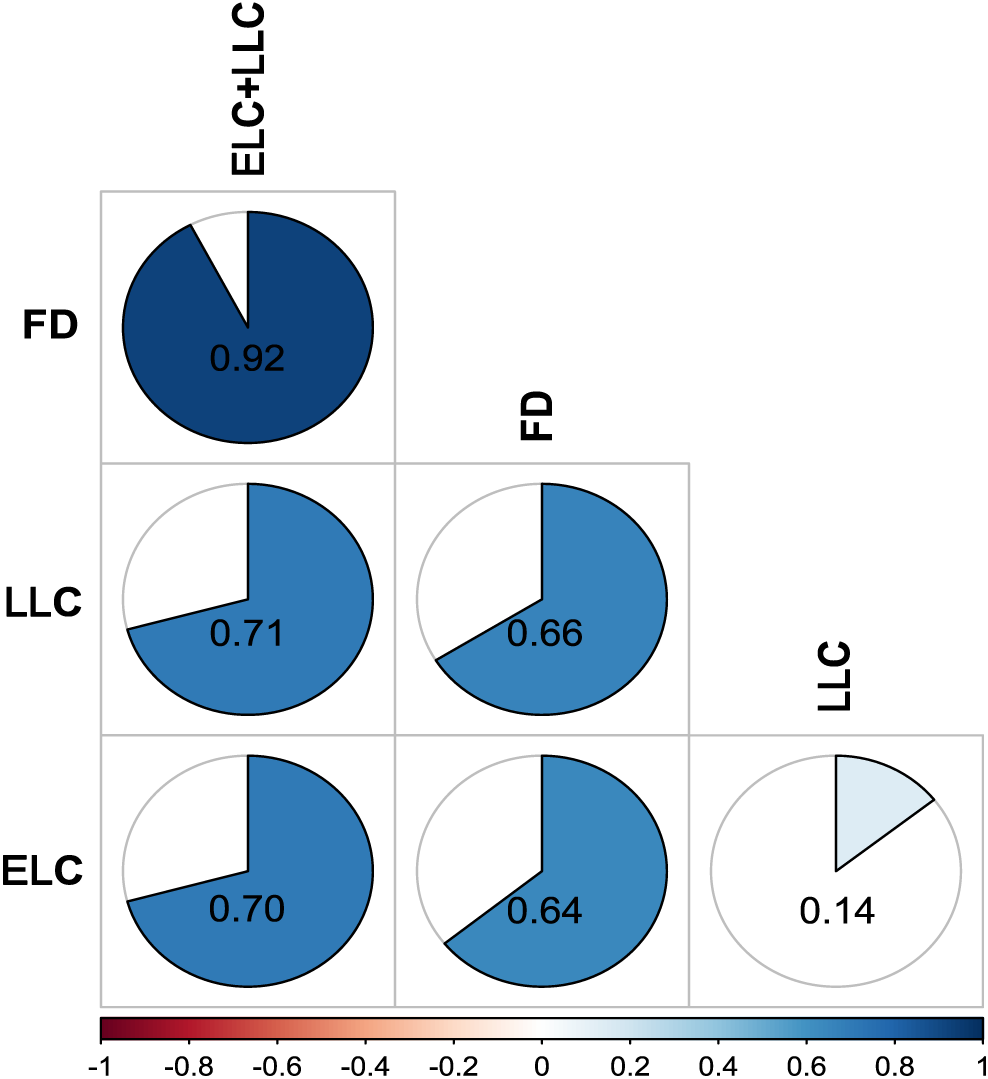
Correlations between disease variables based on means of two years. FD: Final defoliation rating. ELC: Early leaf spot lesion count. LLC: Late leaf spot lesion count. ELC+LLC: Combined lesion counts. Spearman correlation coefficients are indicated within cells.
Accessions differed significantly (P < 0.01) for all disease variables evaluated (Table 1). Final defoliation rating and ELC also indicated significant effects of year (P < 0.05). Variable levels of FD, ELC, and LLC were detected among accessions of the same species. This highlights the importance of selecting germplasm at the accession level rather than species level. Similar findings have been reported in previous studies (Michelotto et al. 2015, Fravero et al. 2009, Pande and Rao, 2001).
With the exception of A. villosa, all perennial/semi-perennial accessions of the A-genome species showed the highest level of resistance with FD values between 1 and 2, and lesion counts lower than 2.5 (Table 1). Among these were A. cardenasii, A. correntina, A. diogoi, A. helodes, A. herzogii, A. kempff-mercadoi, A. kuhlmannii, A. simpsonii, and A. stenosperma. Arachis stenosperma, although described as annual or semi-perennial (Krapovickas et al., 2007), all plants in the field showed a perennial-like growth habit.
Accessions of A. villosa (PI 298636, PI 666104, PI 666106, PI 666108) had moderate levels of resistance to LLS with final defoliation ratings not significantly different from those of the B-genome species A. magna and A. hoehnei. All accessions of A. villosa were susceptible to ELS (Table 1, Fig. 3). The accession from Argentina (PI 298636) was comparatively more resistant than accessions from Uruguay (PI 666104, PI 666106, PI 666108). Plants of this accession showed similarities, but also distinctive morphological characteristics (e.g., pubescence, leaflet shape), suggesting a genetically different germplasm.

Late leaf spot in Arachis kempff-mercadoi. (A) Lesion with spores on the abaxial leaf surface observed under stereomicroscope. (B) Leaflet with lesions. (C) Late season LLS symptoms in field plots.

Early leaf spot in Arachis villosa. (A) Lesion with spores on the adaxial leaf surface observed under stereomicroscope. (B) Early season ELS symptoms. (C) Late season ELS symptoms in field plots.
Among the non-A genome species evaluated, A. hoehnei, A. benensis, and A. magna had the lowest FD levels (2.75-4). Arachis magna has been suggested as a potential candidate for introgression and pyramiding of resistant genes into cultivated peanut (Michelotto et al., 2015). However, further evaluation is needed to select the appropriate A. magna germplasm, as different levels of leaf spot resistance have been reported, and also observed in this study (GKSSc 30093, GKSSc 30097, BRA 33821) for the same accessions when evaluated under different experimental conditions (greenhouse/field inoculation assays) (Subrahmanyan et al., 1985, Favero et al., 2009, Michelotto et al., 2015).
It is suggested that field evaluations are likely to be influenced by canopy microclimates due to the effect of plant architecture on disease development (Coffelt and Porter, 1982; Akem et al. 1992; Chappell et al. 1995). These studies have shown that plants with the upright growth habit and less dense canopy, characteristic of Spanish-type peanut (A. hypogaea ssp. fastigiata) have more resistance to Sclerotinia blight than dense, spreading-type canopy of Virginia-type cultivars (A. hypogaea ssp. hypogaea). However, little is known about the effects of canopy architecture on the development of peanut foliar diseases, or whether the less dense canopy typical of wild Arachis species is more or less conducive for the invasion of some pathogens. Recent studies showed that it was possible to maintain superior disease resistance introgressed from wild Arachis species in a cultivated-type canopy architecture (Leal-Bertioli et al. 2016; Chu et al. 2019). In the present study, all wild species were characterized by a single erect main stem and procumbent lateral branches (Krapovickas and Gregor, 2007). The length, number and branching of lateral branches defined different canopy patterns across species and accessions (Fig. 5). However, despite canopy differences, no apparent correlation was observed between canopy type and disease resistance. For example, accessions of A. batizocoi, characterized by sparse branching, and likely less favorable for fungal invasion, were more susceptible to leaf spots than accessions with dense canopies, such as those of A. cardenasii and A. correntina (Table 1, Fig. 5). Although field experiments were not designed to test the effect of canopy architecture, observations of susceptible and resistant genotypes in adjacent plots indicated that the microclimate created within the canopy of susceptible runner type peanut cultivars did not affect the resistance of wild Arachis genotypes (Fig. 6, A and B). Overall, these observations suggest that canopy architecture may not be a major factor, although this requires further investigations.

Aerial view of the canopy of wild Arachis species in field plots. Newman farm, GA, July 2017.

Resistant and susceptible peanut plants in adjacent plots. (A) A. cardenasii (PI 475997). (B) Susceptible check. White arrows indicate healthy leaves of the resistant A. cardenasii from lateral branches spreading under the canopy of a susceptible check.
Summary and Conclusions
The present study reports the identification of peanut germplasm with resistance to leaf spot diseases which were selected from an environment with high inoculum pressure. As resistance was identified across a diverse Arachis germplasm representing different species, genome type, life cycle, and geographic distribution, it is likely that this resistance encompasses novel genetic sources. Therefore, the highly resistant (nearly immune) accessions of the A-genome species, including A. cardenasii, A. correntina, A. diogoi, A. helodes, A. kuhlmannii, A. kempff-mercadoi, A. simpsonii, and A. stenosperma, along with the moderately resistant accessions of the non-A genome species, including A. benensis (PI 475877), A. hoehnei (PI 681081), and A. magna (PI 468338) could contribute to durable resistance through introgression and gene pyramiding. Further, these novel resources can be leveraged in traditional peanut breeding approaches through the use of marker assisted selection methods to increase introgression efficiency, while minimizing linkage drag.
Acknowledgements
The authors thanks Staci Ingram for technical assistance. This research was supported by USDA-ARS project NP303 6044-42000-011-00D. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Literature Cited
Abdou, Y.A.M., Gregory W.C. and Cooper. W.E. 1974. Sources and nature of resistance to Cercospora arachidicola Hori and Cercosporidium personatum (Berk. et Curtis) Deighton in Arachis species. Peanut Sci. 1: 6– 11.
Akem, C.N., Melouk, H.A. and Smith. O.D. 1992. Field evaluation of peanut genotypes for resistance to Sclerotinia blight. Crop Protect. 11: 345– 348.
Arias, R.S., Sobolev, V.S. Massa, A.N. Orner, V.A. Walk, T.E. Ballard, L.L. Simpson, S.A. Puppala, N., Scheffler, B.E. de Blas, F. and Seijo. J.G. 2018. New tools to screen wild peanut species for aflatoxin accumulation and genetic fingerprinting. BMC Plant Biol. 18: 170.
Chappell, G.F., Shew, B.B. Ferguson, J.M. and Beute. M.K. 1995. Mechanisms of resistance to Sclerotinia minor in selected peanut genotypes. Crop Sci. 35: 692– 696.
Chiteka, Z. A., Gorbet, D.W. Shokes, F.M. Kucharek, T.A. and Knauft. D.A. 1988. Components of resistance to late leafspot in peanut. I. Levels and variability-implications for selection. Peanut Sci. 15: 25– 30.
Chu Y., Chee, P. Culbreath, A. Isleib, T.G. Holbrook, C.C. and Ozias-Akins. P. 2019. Major QTLs for resistance to early and late leaf spot diseases are identified on chromosomes 3 and 5 in peanut (Arachis hypogaea). Front Plant Sci. 10: 883.
Chu, Y., Chee, P. Isleib, T.G. Holbrook, C.C. and Ozias-Akins. P. 2020. Major seed size QTL on chromosome A05 of peanut (Arachis hypogaea) is conserved in the US mini core germplasm collection. Mol. Breeding 40, 6. https://doi.org/10.1007/s11032-019-1082-4.
Coffelt, T.A., and Porter. D.M. 1982. Screening peanuts for resistance to Sclerotinia blight. Plant Dis. 66: 385– 387
Fávero A.P., Simpson, C.E. Valls, J.F.M. Vello. N.A. 2009. Characterization of rust, early and late leaf spot resistance in wild and cultivated peanut germplasm. Sci. Agric. (Piracicaba, Braz.) 66: 110– 117.
Hancock, W.G., Tallury, S.P. Isleib, T.G. Chu, Y. Ozias-Akins, P. and Stalker. H.T. 2019, Introgression analysis and morphological characterization of an Arachis hypogaea × A. diogoi interspecific hybrid derived population. Crop Sci. 59: 640– 649.
Krapovickas, A., and Gregory. W.C. 2007. Taxonomy of the genus Arachis (Leguminosae). Bonplandia 16: 1– 205. (English translation of 1994 Bonplandia 16:1–205 by D.E. Williams and C.E. Simpson).
Leal-Bertioli S.C.M., Godoy, I.J. Santos, J.F. Doyle, J.J. Guimarães, P.M. Abernathy, B.L. Jackson, S.A. Moretzsohn, M.C. Bertioli. D.J. 2018. Segmental allopolyploidy in action: increasing diversity through polyploid hybridization and homoeologous recombination. Am. J. Bot. 105 (6): 1053– 1066.
Michelotto, M.D., Barioni W. de Resende, M.D.V. de Godoy, I.J. Leonardecz, E. et al. 2015. Identification of fungus resistant wild accessions and interspecific hybrids of the genus Arachis. PLOS ONE 10 (6): e0128811.
Pande S., and Rao. R. 2001. Resistance of wild Arachis species to late leaf spot and rust in greenhouse trials. Plant Dis. 85 (8): 851– 855.
R Development Core Team. 2020. R: A language and environment for statistical computing R. Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org/
Simpson, C.E. 2001. Use of wild Arachis species/introgression of genes into A. hypogaea L. Peanut Sci. 28: 114– 116.
Singh, A.K. 1986. Utilization of wild relatives in the genetic improvement of Arachis hypogaea L.-8. Synthetic amphidiploids and their importance in interspecific breeding. Theor. Appl. Genet. 72 (4): 433– 439.
Stalker, H.T. 1984. Utilizing Arachis cardenasii as a source of Cercospora leafspot resistance for peanut improvement. Euphytica 33 (2): 529– 538.
Stalker, H.T., Moss. J.P. 1987. Speciation, cytogenetics, and utilization of Arachis species. Adv Agron. 41: 1– 40.
Stalker, H.T., and Beute. M.K. 1993. Registration of four interspecific peanut germplasm lines resistant to Cercospora arachidicola. Crop Sci. 33: 1117.
Stalker, H.T. 2017. Utilizing wild species for peanut improvement. Crop Sci. 57: 1102– 1120.
Stalker, H.T., Wynne, J.C. and Company. M. 1979. Variation in progenies of an Arachis hypogaea × diploid wild species hybrid. Euphytica 28: 675– 684.
Stalker, H., Beute, M., Shew, B. and Isleib, T. 2002, Registration of five leaf spot-resistant peanut germplasm lines. Crop Sci., 42: 314– 316.
Subrahmanyam, P., Nolt, A.M. Reddy, B.L. Reddy, D.V.R. and McDonald. D. 1985. Resistance to groundnut diseases in wild Arachis species. In: Proceedings of the International Workshop on Cytogenetics of Arachis, Patancheru, India. 31 Oct.-2 Nov. 1983. ICRISAT, Patancheru. pp. 49– 55.
Tallury S.P., Isleib T.G., Stalker H.T. 2009. Comparison of Virginia-type peanut cultivars and interspecific hybrid derived breeding lines for leaf spot resistance, yield, and grade. Peanut Sci. 36: 144– 149.
Tallury, S.P., Hollowell, J.E. Isleib, T.G. and Stalker. H.T. 2014 a. Greenhouse evaluation of section Arachis wild species for Sclerotinia blight and Cylindrocladium black rot resistance. Peanut Sci. 41: 17– 24.
Tallury, S.P., Isleib, T.G. Copeland, S.C. Rosas-Anderson, P. Balota, M. Singh, D. Stalker. H.T. 2014 b. Registration of two multiple disease-resistant peanut germplasm lines derived from Arachis cardenasii Krapov. & Gregory, W.C. GKP 10017. J. Plant Reg. 8: 86– 89.
Wei, T., Simko. V. 2017. R package “corrplot”: Visualization of a correlation matrix (Version 0.84). https://github.com/taiyun/corrplot.
Notes
- USDA-ARS National Peanut Research Laboratory, P.O. Box 509, 1011 Forrester Dr. SE, Dawson, GA 39842. [^]
- USDA-ARS Plant Genetic Resources Conservation Unit, 1109 Experiment Street Griffin, GA 30223-1797. [^]
- North Carolina State University, Headhouse Unit 3 At Method 214, Raleigh, NC. [^] *Corresponding author Email: Alicia.Massa@usda.gov
Author Affiliations