

Introduction
Peanut (Arachis hypogaea L.) is one of the most widely grown legumes by smallholder farmers in Malawi (Nyondo et al., 2018). Peanut is a relatively inexpensive source of dietary protein and other essential nutrients for both urban and rural households (Okello et al., 2010a 2010b). Peanut also serves as an important source of livestock and poultry feed (Kochhar 1986; Usman et al., 2012). However, yield is low (700 kg/ha) in Malawi compared to other countries because most smallholder farmers in Malawi grow peanut with little or no inputs (Ngwira et al., 2019). In addition to low yields, aflatoxin, a mycotoxin produced by Aspergillus flavus and A. parasiticus contaminates peanut-based food products and contributes to poor health (Bowen and Hagan, 2015). Peanut and food products that contain aflatoxin above established levels can limit marketing opportunities (FAO, 2001; Matumba et al., 2015; Waliyar et al., 2010). The European Union and Malawi Bureau of Standards accepts aflatoxin level of 4 μg/kg while the World Health Organization (WHO) standard is 20 μg/kg (Monyo et al., 2012; Otsuki and Wilson, 2001). Aflatoxin contamination adversely affects human health in a number of ways including liver cancer and immunosuppressive effects (Guchi, 2015; WHO, 2006).
Aflatoxin can be present prior to harvest during the growing cycle when peanut is exposed to prolonged high day and night temperatures and drought during pod filling (Payne, 1998; Sanders et al., 1984). Pre-harvest aflatoxin contamination in peanut is often associated with cracked pods that allow entry of soil that contains A. flavus (Craufurd et al., 2006). High relative humidity and soil temperatures ranging from 25 to 35 C are favorable for A. flavus development (Bowen and Hagan, 2015; Cole et al., 1984; Hill et al., 1983). Weather conditions that result in delayed harvest increase the likelihood of greater damage from arthropods and vertebrates that increase pod damage and subsequent movement of soil into pods (Desai et al., 2008). Minimizing stress associated with high temperatures, drought, and damage from other organisms can minimize aflatoxin contamination of peanut going into drying and storing steps in the supply chain (Torres et al., 2014).
Establishing adequate populations of peanut increases yield and uniformity of harvested peanut (Onat et al., 2017; Okello et al., 2010a 2010b). The current recommended density of peanut planted in Malawi is 89,000 plants/ha when established in rows spaced 75 cm apart at a distance of 15 cm between plants in each row (Ngwira et al., 2019). However, Onat et al. (2017) reported that increasing the population to 133,000 plants/ha increased yield compared with lower densities in Turkey. Greater yields were also noted in the United States (Kvien et al., 1987) and India (Rasekh et al., 2010) when higher plant populations were established as compared with lower plant populations. As farmers consider increasing seeding rates to achieve higher plant populations, it is important to determine if this approach affects aflatoxin contamination. The impact of plant population on aflatoxin contamination has not been documented in the peer-reviewed literature.
Harvesting peanut when the kernels and pods reach physiological maturity often results in greater yields, higher market grade characteristics (Okello et al., 2010a 2010b), and can minimize seed infection by A. flavus (Mehan et al., 1986; Sanders et al., 1985). Harvesting peanut prior to physiological maturity can result in a distribution of kernels that contain a higher percentage of smaller and less mature kernels as compared with harvesting at physiological maturity (Carter et al., 2017). When peanut is harvested past physiological maturity, pods have greater exposure to pathogens and arthropods that can cause damage (Okello et al., 2010a 2010b; Singh and Oswalt, 1995). Pods that are past physiological maturity for an extended period of time, especially if soil moisture is adequate, can sprout and result in lower yield and quality (Nautiyal et al., 2001; Singh and Oswalt, 1995). Pods can also shed from plants and may not be harvestable (Kaba, 2014).
The majority of farmers in Malawi establish plant populations lower than what is needed to optimize yield because of seed quality issues and expense (Ngwira et al., 2019). Farmers also do not fully appreciate the relationship of harvest date, physiological maturity, and the impact of pests on yield and quality, including the human health and marketing ramifications of contamination with aflatoxin (Kaba, 2014). The relationship between plant population and harvest date has not been evaluated in Malawi in a systematic way with respect to peanut yield and aflatoxin contamination. Therefore, research was conducted to determine the impact of plant population and harvest date on peanut yield and contamination by aflatoxin at harvest in Malawi.
Materials and Methods
The experiment was conducted at Mpatsanjoka farm in Salima district of Malawi (13°42.740′S, 034°28.879′E) during crop cycles in 2015-2016 and 2016-2017. Mpatsanjoka farm is located near Lake Malawi at an elevation of 550 m above sea level. Monthly average rainfall from Nov through March for each cropping cycle is presented in Table 1. No measurable rainfall was observed after March of each year. Soil was a sandy loam with pH 5 and organic matter content of 2.7%. Peanut was seeded on 13 December 2015 and 22 December 2016 in raised seedbeds with a height of 20 to 30 cm (Table 2). Immediately after planting, dimethenamid-P (Frontier-P herbicide, Australian Pesticides and Veterinary Medicine Authority, Kingston, Australia) was applied at 0.84 kg ai/ha to control weeds. Hand weeding was used to throughout the season to minimize weed interference. No other pesticides were applied to control arthropods or pathogens.

Monthly rainfall recorded at Mpatsanjoka farm in Salima district of Malawi during cropping cycles of 2015-2016 and 2016-2017 from Nov through March. No measurable rainfall occurred after March during the peanut growing cycle.
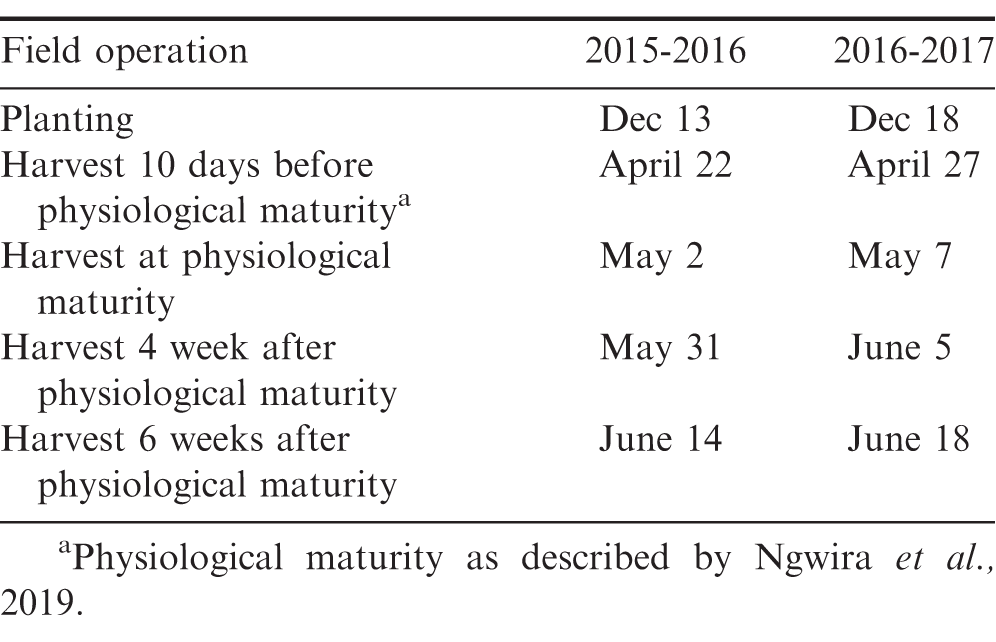
Planting and harvest dates for peanut during two growing cycles in Malawi.
Treatments consisted of the virginia market type cultivar CG7 (Ngwira et al., 2019) seeded in single, twin, and triple row patterns at rates of 89,000, 178,000, and 285,000 seed/ha to establish low, medium, and high plant populations, respectively (Table 3). The seeding rate of 89,000 seeds/ha was established on beds spaced 75 cm apart with one row of peanut with 15 cm between seeds. The seeding rate of 178,000 seeds/ha was established on beds with 75-cm centers with two rows of peanut spaced 18 cm apart with 15 cm between seeds. The seeding rate of 285,000 plants/ha was established on beds with 75-cm centers with three rows of peanut spaced at 18 cm with 7 cm between seeds. The final in-row spacings for these respective plant populations/ha during each cycle are presented in Table 3. Seeds were treated with thiram (Thiram 50WP, Bayer Crop Science, Research Triangle Park, NC) at 3g/kg of peanut seed and planted in furrows created manually at a depth of 3 cm. Within each plant population, peanut was harvested 10 d before physiological maturity, at physiological maturity, 4 wk after physiological maturity, and 6 wk after physiological maturity. The internal color of pods was used to determine maturity by collecting representative plants across the field. When 70% of pods visible darkening caused by removal of the endocarp as kernels increased in size and development, peanut was considered physiologically mature (Ngwira et al., 2019). Approximately 50% of pods expressed this level of darkening 10 d prior to physiological maturity. After gently removing peanut pods from the soil, above-ground vegetation, pods, and remaining roots were placed on a Mandela cock (Ngwira et al., 2019). When pod moisture was 8.5%, total plant biomass, pod weight, grain weight, and aflatoxin contamination in pods were determined.

Planting pattern, seeding rate, and average plant population at harvest during two growing cycles in Malawi.
Aflatoxin concentration was determined using 300 g of shelled peanut from each plot. The sample was weighed and homogenized using a hand grinder (Globe Trek, Navi Mumbai, Mumbai) and thoroughly mixed. A 10-g sub-sample was removed and agitated in 30 ml of ethanol (65%) and water (35%) for 1 min. The sub-sample was filtered using a 500 μL pipette (Fisher Scientific, Pittsburg, PA) with the diluent. One hundred μL of the filtered liquid was inserted into the sampling cup and left in the cup for 6 min. Neogen Reveal Q+ lateral flow strips (Neogen Corp., Lansing, MI) were inserted into the strip holder (Neogen Corp., Lansing, MI) to determine aflatoxin concentration using the Mobile Assay mReader software (Mobile Assay Inc., Boulder, CO).
Data for total biomass, pod weight, grain weight, and aflatoxin contamination were subjected to analysis of variance for the split-plot design using GENSTAT 18th edition computer package (VSN international, England, United Kingdom). Seeding density served as the whole plot unit and harvest date served as the sub-plot unit. Differences of main effects and interactions were separated using Fisher’s Protected LSD test at p < 0.05.
Results and Discussion
Interactions of growing cycle by seeding rate by harvest date were not significant for plant biomass at harvest, pod yield, and aflatoxin concentration (p > 0.05). However, main effects of seeding rate and harvest date, the interaction of growing cycle and seeding rate, and the interaction of growing cycle and harvest date were significant. Data for plant biomass, pod yield, and aflatoxin concentration will be presented for these interactions combined over growing cycle and the other treatment factor (Tables 4 and 5). Oakes et al. (2020) reported that both seeding rate and harvest date affected pod yield but response to these treatment factors was independent.
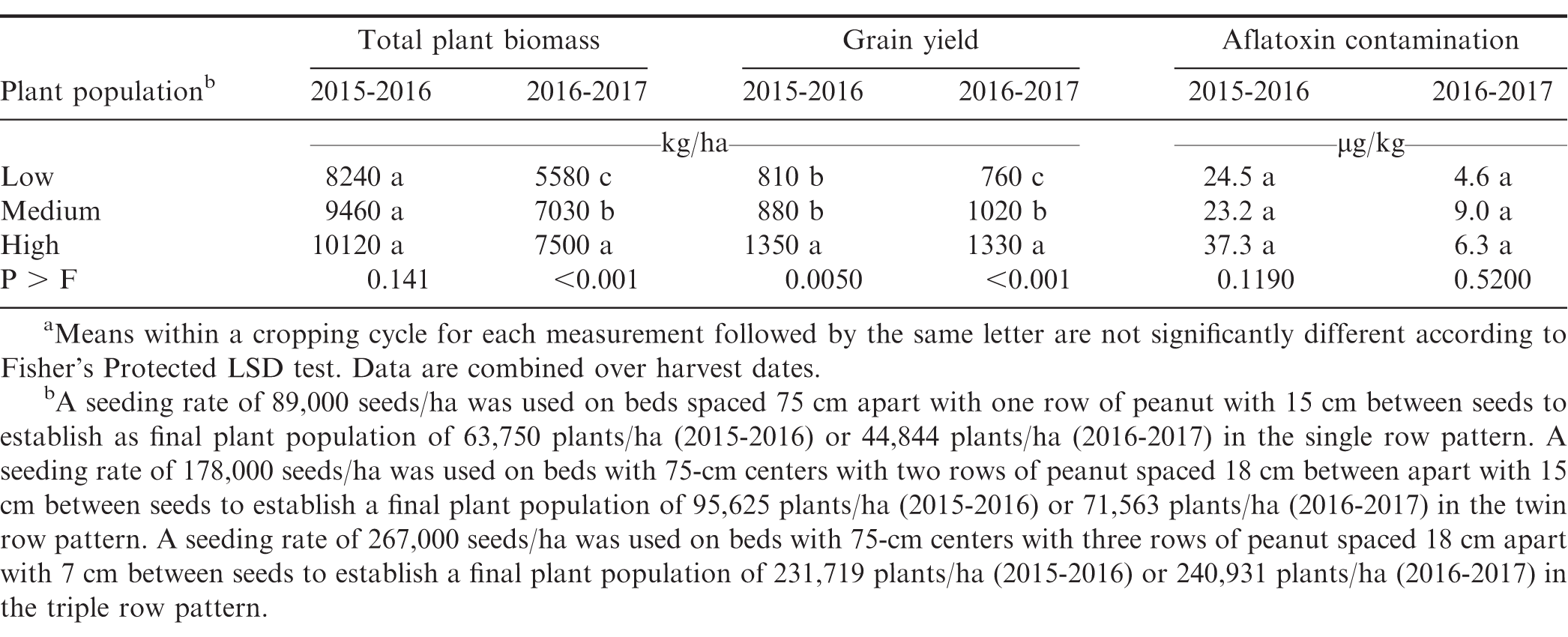
Influence of seeding rate on total peanut biomass, peanut grain yield, and aflatoxin contamination.a
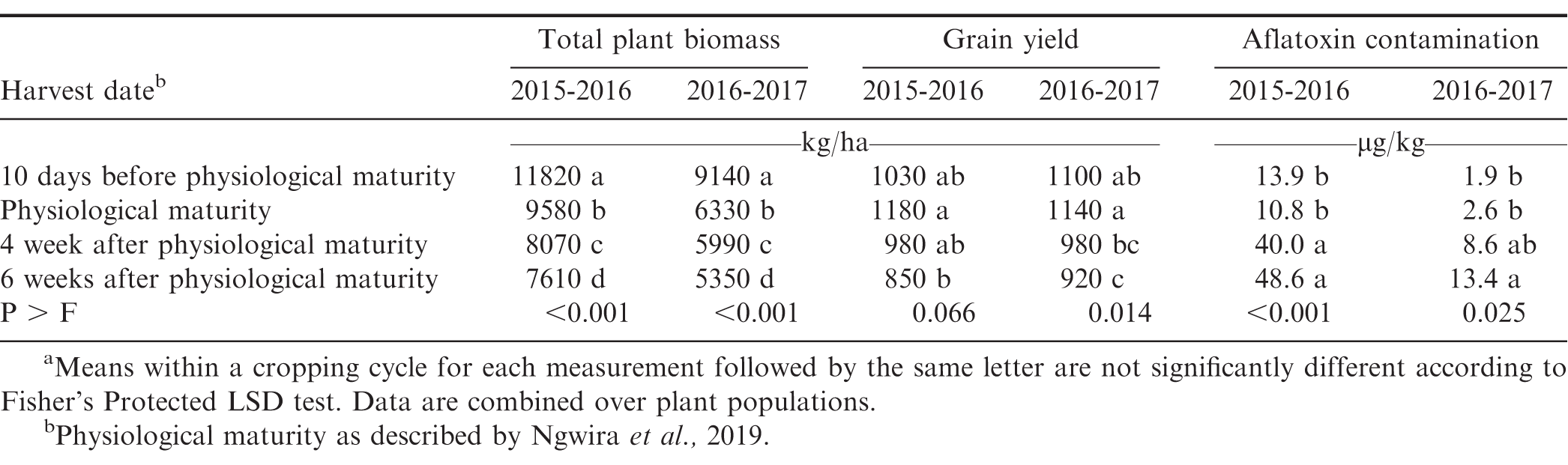
Influence of harvest date on total peanut biomass, peanut grain yield, and aflatoxin contamination.a
Above-ground plant biomass increased as seeding rate and subsequent plant population increased in one of the two growing cycles (Table 4). In 2016-2017 growing cycle, plant biomass increased for each increase in plant density. Bell et al., (1987) reported that peanut biomass increased from 12,600 kg/ha to 16,900 kg/ha with increasing plant density up to the maximum density of 588,000 plants/ha.
Peanut pod yield was greater in both growing cycles for the seeding rate of 285,000 seed/ha compared with the lower seeding rates (Table 4). In the 2015-2016 growing cycle, no difference in yield was observed for the low and medium seeding rates. However, in the 2016-2017 growing cycle, peanut yield for the medium seeding rate exceeded that of the low seeding rate. Aflatoxin contamination at harvest was not affected by seeding rate in either growing cycle.
Increasing the seeding rate often ensures greater uniformity of pod maturity, improved quality of grain for marketing, and maximum yield (Okello et al., 2010a 2010b). However, variation in response to seed spacing has been observed. Konlan et al. (2013) in Ghana reported that decreasing spacing between seeds increased yield by 6.2% in one years and 16.0% in a second year of the study. Rasekh et al. (2010) reported higher yields when the in-row seeding rate was increased from 3 plant/m to 8.3 plants/m. Awal and Aktar (2015) and Gabisa et al. (2017) reported greater yield with a plant population of was increased. El Naim et al. (2011) reported that peanut plants spaced 10 cm apart yielded 40% less than peanut planted 40 cm apart under rain-fed conditions.
Above-ground peanut biomass was greater 10 d prior to physiological maturity compared to mass at physiological maturity (Table 5). When harvest was delayed past physiological maturity, biomass continued to decrease. This was due to significant vegetative growth and by this time the crop did not lose its leaves while the soil was still moist. Peanut pod yield was similar when harvested at physiological maturity, or 10 d prior to physiological maturity (Table 5). In the first growing cycle, yield was similar when peanut was harvested at physiological maturity or 4 wk after physiological maturity. In contrast, in the second growing cycle, peanut yield was lower when harvested 4 wk after physiological maturity than harvest at physiological maturity. Delaying harvest to 6 wk after physiological maturity resulted in the lowest grain yield. The decrease in pod yield with the delayed harvesting was due to field losses as many remaining in the field due to natural pod shed. Young et al. (1982) estimated typical digging losses of 8% of total yield, but can reach 40% at dates when harvest is delayed past optimal maturity. Okello et al. (2010a 2010b) reported that delayed harvesting causes yield losses of greater than 400 kg/ha and kernel quality reduced by 3%.
Aflatoxin contamination was similar when peanut was harvested 10 d prior to physiological maturity or at physiological maturity (Table 5). In the first growing cycle, aflatoxin contamination was greater when harvest was delayed by 4 and 6 wk past physiological maturity compared with harvest 10 d prior to physiological maturity or at physiological maturity. By 4 and 6 wk after physiological maturity, aflatoxin contamination was 40.0 to 48.6 μg/kg. During the second growing cycle, aflatoxin contamination did not exceed 13.6 μ/kg. The difference in aflatoxin due to harvest date most likely was associated with rainfall patterns during the latter part of the growing cycle in March. Rainfall ceased during the 2015-2016 growing cycle in March while rainfall during March 2017 was during 264 mm and just under twice the total rainfall during the growing cycle (Table 1). Adequate rainfall which reduces aflatoxin contamination in peanut because it increases tissue integrity hence reduce invasion of A. flavus (Diao et al., 2015). The lower concentration of aflatoxin most likely was associated with complete maturation of pods when rainfall was more abundant in March which minimized infection when soil moisture in soil was higher. Sanders et al. (1984) reported that peanut pods developed without damage to shells and limit entry of Aspergillus spp. to colonize pods. Okello et al. (2010a 2010b) reported that harvesting peanut at optimum maturity reduces incidences of aflatoxin contamination. Peanut harvested after physiological maturity had high aflatoxin contamination because of over maturity and delayed harvesting which increases aflatoxin contamination (Diener and Davis, 1977; Okello et al., 2010a 2010b).
In summary, increasing seeding rates and subsequent plant populations up to 285,000 seed/ha resulted in the greatest yield across the two growing cycles. However, seeding rate had no effect on aflatoxin contamination at harvest. It is postulated that higher plant populations likely shade soil and create cooler soil environment that is less conducive to A. flavus growth and infection. However, our results did not support that hypothesis. It was also postulated that harvesting peanut prior to physiological maturity would result in less aflatoxin compared with harvesting at physiological maturity or past physiological maturity. Our results are in contrast to those by Bowen and Hagan (2015) who reported less aflatoxin when peanut was harvested earlier than recommended to optimize pod yield when conditions were favorable for A. flavus development. However, our results were consistent with those of Young et al., (1982) demonstrating that delaying harvest past physiological maturity can result in lower yield due to leaf and pod shed due to disease and natural processes and sprouting of seed or damage caused by arthropods in soil. Pod damage caused by arthropods can allow soil to enter pods and increase the amount of A. flavus in pods and subsequently cause greater aflatoxin contamination.
This is the first experiment in the peer-reviewed literature that has addressed the impact of seeding rate and subsequent contamination by aflatoxin. Our results indicate that plant population and harvest date do not interact for pod yield or aflatoxin contamination. Oakes et al. (2020) also reported that both seeding rate and harvest date affected peanut pod yield independently. It is important to note that plant populations were established using three different planting patterns with peanut plants distributed differently across beds. Additional research is needed to determine the impact of a more dense population using a similar planting pattern to determine the impact of plant population on yield and aflatoxin contamination.
Acknowledgements
This publication was made possible through support provided by the Office of Agriculture, Research and Policy, Bureau of Food Security, U.S. Agency for International Development, under the terms of Award No. AID-ECG-A-00-07-0001 to The University of Georgia as management entity for U.S. Feed the Future Innovation Lab on Peanut Productivity and Mycotoxin Control (2012-2017). The opinions expressed herein are those of the authors and do not necessarily reflect the views of the U.S. Agency for International Development. Appreciation is expressed to technical staff and farmers for assistance with this research. Appreciation is expressed to technical support at the Bunda College Campus.
Literature Cited
Awal MAand Aktar L. 2015. Effect of row spacing on the growth and yield of peanut (Arachishypogaea L. ) stands. International Journal of Agriculture, Forestry and Fisheries, 3: 7– 11. Available at: http://openscienceonline.com/journal/archive2?journalId=706&paperId=1365 [Accessed 12 Feb 2021]
Bell, M. J., Muchow, R. C. and Wilson. G. L. 1987. The effect of plant population on peanuts (Arachis hypogaea) in a monsoonal tropical environment. Field Crops Research. 17: 91– 107.
Bowen, K. L. and Hagan. A. K. 2015. Temperature and moisture conditions that affect aflatoxin contamination of peanuts. Peanut Sci. 42: 121– 127.
Carter, E. T, Rowland, D. L. Tillman, B. L. Erickson, J. E. Grey, T. L. Gillett-Kaufman, J. L. and Clark. M. W. 2017. Pod maturity in the shelling process. Peanut Sci. 44: 26– 34.
Cole, R. J., Blankenship, P. D. Hill, R. A. and Sanders. T. H. 1984. Mean geocarposphere temperature that induce preharvest aflatoxin contamination of peanuts under drought stress. Mycopathologia. 91: 41– 46. Available at: https://ncbi.nlm.nih.gov/pubmed/3930968. [Accessed 12 Feb 2021]
Craufurd, P. Q, Prasad, P. V. V. Waliyar, F. and Taheri. A. 2006. Drought, pod yield, pre-harvest Aspergillus infection and aflatoxin contamination on peanut in Niger. Field Crops Research. 98: 20– 29.
Desai S, Khandar, R. R. Waliyar, F. Thakur, R. P. Dhruj, I. U. Nigam, S. N. and Bandyopadhyay. A. 2005. A Hazard Analysis Critical Control Point (HACCP) based approach for an integrated management of aflatoxin contamination in groundnut in Gujarat, India, In: Proceedings of International Peanut Conference: Prospects and Emerging Opportunities for Peanut Quality and Utilization Technology A9-15. Thailand: Kasetsart University, Bangkok, 102. Available at: http://oar.icrisat.org/5409/1/IPC_102_%202005.pdf [Accessed 12 Feb 2021]
Diao, E., Dong, H. Hou, H. Zhang, Z. Ji, N., and Ma. W. 2015. Factors influencing aflatoxin contamination in before and after harvest peanuts: A Review. J. of Food Research. 4: 148– 154. Available at: http://doi.org/10.5539/jfr.v4n1p148 [Accessed 12 Feb 2021]
Diener, U. L. and Davis. N. D. 1977. Aflatoxin formation in peanut by Aspergillus flavus. Ala. Agric. Exper. Sta. Bull. No. 493, Auburn University. Available at: https://pdfs.semanticscholar.org/1e40/a28324fb6ca138e8d267898e64e0fced722e.pdf. [Accessed 12 Feb 2021]
El Naimp, A. M., Eldouma, M. A. Ibrahim, E. A. and Moayad Zaied MB M. B. . 2011. Influence of plant spacing and weeds on growth and yield of Peanut (Arachis hypogaea L) in rain-fed of Sudan. Advances in Life Sciences. 1: 45– 48. DOI: 10.5923/j.als.20110102.08.
FAO [Food and Agriculture Organization]. 2001. Manual on the application of HACCP system in mycotoxins prevention and control. FAO Food and Nutrition Paper No. 73. FAO, Rome, Italy. ISSN 0254-4725.
Gabisa, M., Tana, T. and Urage. E. 2017. Effect of planting density on yield components and yield of Groundnut (Arachis hypogaea L.) varieties at Abeya, Borena Zone Southern Ethiopia. International Journal of Scientific Engineering and Applied Science. 3: 2395– 3470. Available at: http://ijseas.com/volume3/v3i3/ijseas20170317.pdf [Accessed 12 Feb 2021]
Guchi, E. 2015. Aflatoxin contamination in groundnut (Arachis hypogaea L. ) caused by Aspergillus species in Ethiopia. J. Applied and Environmental Microbiology. 3: 11– 19. Available at: file:///C:/Users/dljorda2/Downloads/EphremsPublishedV.3PhD.pdf. [Accessed 12 Feb 2021]
Hill, R. A., Blankenship, P. D. Cole, R. J. and Sanders. T. H. 1983. Effects of soil moisture and temperature on preharvest invasion of peanuts by the Aspergillus flavus group and subsequent aflatoxin development. Applied and Environmental Microbiology. 45: 628– 633. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC242335/pdf/aem00171-0286.pdf [Accessed 12 Feb 2021]
Kaba, J. S., Ofori, K. and Kumaga F. K. . 2014. Inter-relationships of yield and components of yield at different stages of maturity in three groundnut (Arachis hypogaea L) varieties. International Journal of Life Sciences. 2: 43– 48.
Konlan, S., Sarkodie-Addo, J. Asare, E. and Kombiok. M. J. 2013. Groundnut (Arachis hypogaea L.) varietal response to spacing in the Guinea savanna agro-ecological zone of Ghana: Growth and yield. African Journal of Agricultural Research. 8: 2769– 2777. DOI: 10.5897/AJAR12.1313.
Kvien, C. S. and Bergmark. C. L. 1987. Growth and development of the Florunner peanut cultivar as influenced by population, planting date and water availability. Peanut Sci. 14: 11– 16.
Matumba, L., Van Pouche, C. Monjerezi, M. Ediage, E. N. and De Saeger. S. 2015. Concentrating aflatoxins on the domestic market through groundnut export: A focus on Malawian groundnut value and supply chain. Food Control. 51: 236– 239. DOI: 10.1016/j.foodcont.2014.11.035.
Monyo, E. S., Njoroge, S. M. C. Coe, R., Osiru, M. Madinda, F. Waliyar, F. and Anitha. S. 2012. Occurrence and distribution of aflatoxin contamination in groundnuts (Arachis hypogaea L) and population density of aflatoxigenic Aspergilli in Malawi. Crop Protection. 42: 149– 155.
Nautiyal, P. C, Bandyopadhyay, A. and Zala. P. V. 2001. In situ sprouting and regulation of fresh seed dormancy in Spanish type groundnut (Arachis hypogaea). Field Crop Research. 70: 233– 241. DOI: 10.1016/S0378-4290(01)00143-5.
Ngwira, A., Madzonga, O. Chunga, P. Z. Siyeni, D. Chintu, J. M. M. Simwaka, P. and Yohane. E. 2019. Guide for groundnut, pigeon pea, sorghum and finger millet production in Malawi. Available at: https://www.researchgate.net/publication/336564876. [Accessed 12 Feb 2021]
Nyondo, C. J., Nankhuni, F. J. and Me-Nsope. N. 2018. Systematic analysis of groundnut production, processing and marketing in Malawi. Policy Research Brief 64. Feed the Future Innovation Lab for Food Security Policy. Available at: https://www.canr.msu.edu/fsp/publications/policy-research-briefs/policy_brief_64.pdf. [Accessed 12 Feb 2021]
Oakes, J. C. Balota, M. Jordan, D. L. Hare, A. T. and Sadeghpour. A. 2020. Peanut response to seeding density and digging date in the Virginia-Carolina region. Peanut Sci. 47: 180– 188.
Okello, D. K., Biruma, M. and Deom. C. M. 2010 a. Overview of groundnuts research in Uganda: Past, present and future. African Journal of Biotechnology. 9: 6448– 6459. DOI: 10.5897/AJB09.013.
Okello, D. K., Kaaya, A. N. Bisikwa, J. Were, M. and Oloka. H. K. 2010 b. Management of aflatoxins in groundnuts: A manual for farmers, processors, traders and consumers in Uganda. National Agricultural Research Organisation, Entebbe. Available at: https://www.researchgate.net/profile/David_Kalule/publication/260247426_Management_of_Aflatoxins_in_Groundnuts_A_manual_for_Farmers_Processors_and_Traders_in_Uganda/links/5900d93d4585156502a04941/Management-of-Aflatoxins-in-Groundnuts-A-manual-for-Farmers-Processors-and-Traders-in-Uganda.pdf. [Accessed 12 Feb 2021]
Onat, B., Bakal, H. Gulluoglu, L. and Arioglu. H. 2017. The effects of row spacing and plant density on yield and yield components of peanut grown as a double crop in Mediterranean environment in Turkey. Turkish Journal of Field Crops. 22: 71– 80. DOI: 10.17557/Tjfc.303885.
Otsuki, T. and Wilson. J. S. 2001. Saving two in a billion: A case study to quantify the trade effect of European food safety standards on African exports. Food Policy. 26: 495– 514. Available at: https://www.sciencedirect.com/science/article/abs/pii/S0306919201000185. [Accessed 12 Feb 2021]
Payne, G. A. 1998. Process of contamination by aflatoxin producing fungi and their impact on crops. Pages 279– 300 In Sinha K. and Bhatnagar D. (eds.). Mycotoxins in Agriculture and Food Safety. New York: Marcel Decker.
Rasekh, H., Asghari, Safarzadeh J. M. N. Wishkai, S. L., Massoumi, R. Zakerinejad. 2010. Effect of planting pattern and plant density on physiological characteristics and yield of peanut (Arachis hypogea L.) in Iran. Research Journal of Biological Sciences. 5: 542– 547. Available at: http://www.medwelljournals.com/abstract/?doi=rjbsci.2010.542.547. [Accessed 12 Feb 2021]
Sanders, T. H., Blankenship, P. D. Cole, R. J. and Hill. R. A. 1984. Effect of soil temperature and drought on peanut pod and stem temperahires relative to Aspergillusjlaous invasion and aflatoxin contamination. Mycopathologia. 86: 51– . 54. Available at: https://ncbi.nlm.nih.gov/pubmed/6429541. [Accessed 12 Feb 2021]
Sanders, T.H. Cole, R. J. Blankenship, P. D. and Hill. R. A. 1985. Relation of environmental stress suration to Aspergillus flavus invasion and aflatoxin production in preharvest peanuts. Peanut Sci. 12: 90– 93.
Singh, Fand Oswalt. D. L. 1995. Groundnut production practices. ICRISAT Skill Development Series No. 3. ICRISAT Training Program. Pradesh, India. Available at: http://gis4agricgh.net/POLICIES/Groundnut%20Production.pdf. [Accessed 12 Feb 2021]
Torres, A. M., Barros, G. G. Palacios, S. A. Chulze, S. N. and Battilani. P. 2014. Review on preand post-harvest management of peanuts to minimize aflatoxin contamination. Food Research International. 62: 11– 19.
Usman, I., Taiwo, A. Haratu, D. and Abubakar. M. 2012. Socio-economic factors affecting groundnut production in sabongari local government of Kaduna State, Nigeria. International Journal of Food and Agricultural Economics. 1: 41– 48. http://ageconsearch.umn.edu/record/156141/files/41.pdf?version=1 [Accessed 12 Feb 2021]
Waliyar, F., Osiru, M. Siambi, M. and Chinyamunyamu. B. 2010. Assessing occurrence and distribution of aflatoxins in Malawi. ICRISAT Final Report Grant No. 08598. Available at: http://oar.icrisat.org/7380/1/Aflatoxins.pdf. [Accessed on 12 Feb 2021]
WHO [World Health Organization]. 2006. Street food vending in the region: food safety challenges. AFRO Food Safety Newsletter - World Health Organization - Food Safety Unit (FOS). 2: 5– 8. Available at: https://afrolib.afro.who.int/documents/2006/eng/FOSNewsletter2_5_8.pdf. [Accessed 12 Feb 2021]
Young, J. H., Person, N. K. Donald, J. O. and Mayfield. W. D. 1982. Harvesting, curing and energy utilization. Pages 458– 485 In Pattee H.E. and Young C. T. (eds). Peanut Science and Technology. Am. Peanut Res. Educ. Society Inc., Yoakum. TX, USA.
Notes
- Crop and Soil Sciences Department, Lilongwe University of Agriculture and Natural Resources, Bunda Campus, Lilongwe, Malawi; [^]
- Exagris Africa Limited, Lilongwe, Malawi; [^]
- Department of Entomology and Plant Pathology, North Carolina State University, Raleigh, NC; [^]
- Department of Crop and Soil Sciences, North Carolina State University, Raleigh, NC. [^] *Corresponding author Email: david_jordan@ncsu.edu
Author Affiliations