

Introduction
The global population is on a steady increase with the expectation that it will peak at approximately 9.5 billion people by 2050 (Godfray et al., 2010). This imposes a major challenge of ensuring food supply in sufficient quantities and quality. The problem is compounded by the effects of climate change and economic as well as political volatility that have stymied progress towards achieving food security (Lobell et al., 2011; Dawson et al., 2016). In the past, addressing these challenges involved intensifying agricultural production with the aim of meeting calorie demands as exemplified in the green revolution (Pingali, 2012; Mehta, 2018). However, it is increasingly recognized that ensuring food security necessitates looking beyond hunger and facilitating access to nutritious food in adequate quantities (FAO, 2019).
The nutritional profile of peanut makes it an excellent source of both caloric components and micronutrients that exert a positive contribution to human health. The peanut seed is composed of 41.2 to 58.6% oil, 12 to 36% protein (Savage and Keenan, 1994) as well as a host of other beneficial micronutrients such as tocopherols, folates, flavonoids, phenolics and free amino acids (Francisco and Resurreccion, 2008). The compendium of nutrients and bioactive compounds makes peanut a useful nutraceutical (Akram et al., 2018; Toomer, 2018). In this regard, it has been extensively used as a ready to use therapeutic food (RUTF) in treating and reversing the effects of severe acute malnutrition especially in children (Bailey, 1963; Enserink, 2008; Israëls et al., 2009). It is also used as a ready to use supplementary food (RUSF) in managing early stage malnutrition (Variath and Janila, 2017).
The oil profile of the legume contains a proportionally favorable composition of fatty acids. Up to 90% of the fatty acid content is comprised of palmitic, oleic and linoleic acid, with oleic and linoleic acid making up 80% of this composition (Norden et al., 1987; Dean et al., 2009). In high oleic peanut cultivars, the ratio of oleic acid to linoleic acid (O/L) makes peanut an ideal source of monounsaturated fatty acids, which have been shown to have good health benefits. These include reduction of hypertensive effects (Terés et al., 2008), anti-inflammatory effects (Carrillo et al., 2012) as well as alleviation of type II diabetes and reversal of negative effects of obesity (Vassiliou et al., 2009). In addition, the high oleic peanut has been demonstrated to reduce the levels of serum cholesterol and low-density lipoprotein cholesterol, resulting in improvement of serum lipid and apolipoprotein levels in postmenopausal women (O’Byrne et al., 1997). When used as feed, high oleic peanut increases the amount of monounsaturated fat by up to 32% while reducing the amount of polyunsaturated fat in pigs (Myer et al., 1992). This indirectly contributes to human nutrition. Aside from its health consequences, this oil composition confers oxidative stability, with a net positive effect on the quality of its products (O’Keefe et al., 1993).
China produced 36% of global peanut in 2017, followed by India, US and Nigeria each of which accounted for 19, 7 and 5% of global production, respectively (FAOSTAT, 2017). In China and India, most of the peanut produced is crushed for oil extraction with high oil content varieties being preferred. In the US the main use is confection and other food purposes with the preference being for lower oil content. In Africa, food and oil use of peanut is comparable (Birthal et al., 2010; FAOSTAT, 2014; Janila et al., 2016a; 2016b). This varied use of the legume underscores the need for varieties with both good nutritive and oil profiles.
Since the discovery of two high oleic lines (F435-2-1 and F435-2-2) with 80% oleic and 2% linoleic acid (Norden et al., 1987), incorporating the high O/L trait in breeding lines has become integrated in the breeding programs of the US. This trait results from the action of two non-allelic homeologous genes, ahFAD2A and ahFAD2B, with 99% sequence homology. Inactivation of these genes results in absence of Δ12-desaturase enzyme activity that catalyzes conversion of oleic acid to linoleic acid (Jung et al., 2000a; 2000b). In the US, development of high O/L cultivars was accelerated by implementing the molecular marker for the mutant allele of ahFAD2B in breeding programs, enabling the pyramiding of the high O/L trait with other traits of interests (Chu et al., 2009,2011). Consequently, marker assisted selection is routinely implemented to pyramid the high O/L trait with other traits of interest (Chu et al., 2011). Other peanut producing countries such as India, China, Brazil, Argentina, South Africa, Israel, Japan and Australia are also including the high O/L trait in their peanut (Janila et al., 2016b; Nawade et al., 2018). However, information on studies of oil and other seed composition traits from West African lines is seldom available.
In this study, we analyzed the seed composition profile of a chromosome segment substitution line (CSSL) population. The cultivated parent used to create the population was Fleur 11, an important spanish type peanut cultivar grown in Senegal, West Africa. In a previous study, Fonceka et al. (2012) crossed this variety with a synthetic allotetraploid peanut derived from the diploid progenitors of cultivated peanut (Fávero et al., 2006). A total of 122 lines were developed, each of which contained a small chromosomal segment from the wild in such a way that the entire wild genome was represented in the whole CSSL population (Fonceka et al., 2012). Since the CSSL population has a uniform genetic background, it was hypothesized that phenotypic variations that distinguish the lines from Fleur 11 would be due to presence of wild introgressions. With that in mind, our objective was to investigate the effects of these introgressions on oil content, fatty acid composition, sugar profile and protein content on a subset of the CSSLs relative to Fleur 11.
Materials and Methods
The composition profiles of 77 CSSLs were assayed in the growing seasons of 2016 and 2017. The parental check Fleur 11, was included in both years while two other cultivated checks, OLin (Simpson et al., 2003) and New Mexico Valencia A (Hsi and Finkner, 1972), were planted in the second year. The plants were grown in the field at the University of Georgia (Tifton) Gibbs farm (31°26′04.7” N 83°35′18.4” W). The soil type was Tift loamy sand (fine-loamy, kaolinitic, thermic Plinthic Kandiudults), and the trial plan was a randomized complete block design (RCBD) with three replicates in each year. Each two-row plot was 3 m long with the seeds planted at a spacing of 5 cm. Conventional agronomic practices included scheduled fungicide and pesticide applications as well as regular irrigation as required. At the end of the season, seeds for the population were harvested, dried and inspected for maturity as indicated by darkening of the endocarp. From each line, 5 grams of fully mature seed were collected, packaged in a labeled freezer bag and shipped to the USDA ARS Market Quality and Handling Research unit (MQHRU) lab at the North Carolina State University. Assays for fatty acid profile, sugar profile, total fat and total protein were conducted as follows.
Total protein was determined using a Sprint Titrator (CEM, Matthews, NC). This instrument uses a proprietary solution to tag the amine side chains of proteins. The resulting solution is fed through a visible light detector that converts the signal to percent protein. The instrument was factory calibrated for peanuts.
Total sugars were determined using the method first described by Pattee et al. (2000a). Mono, di and trisaccharides were extracted from the samples after defatting with hexane. A mixture of chloroform, methanol and water (60/25/15 v/v/v) was used. The organic solvents were removed from the extracts by evaporation under nitrogen. The remaining aqueous solution was spiked with internal standard solution (lactose and cellobiose, Sigma Chemical Corp., St. Louis, MO). Finally, 50 μL of each sample solution was diluted to 2 mL with water, passed through a Dionex OnGuard-H filter (Dionex, Sunnyvale, CA), and injected onto a Dionex Bio LC system. A Dionex CarboPac™ PA-1 column (250 mm length, 4 mm i.d.) and a Pulsed Amperometric Detector (PAD) was used. The column was heated to 25°C. The mobile phase was 200 mM sodium hydroxide at a flow rate of 1.0 mL/min. The sugars present were quantified using response ratios to the internal standards compared to those of authentic standards of myo-inositol, glucose, fructose, sucrose, raffinose, and stachyose (Sigma).
The total fat in the samples was determined using a Mini Spec Seed Analyzer, (Bruker Instruments, Billerica, MA). Five grams of sample were loaded into a glass sample tube and inserted into the instrument. This instrument uses time domain or low power nuclear magnetic resonance. It is used for total fat determination in oilseeds according to AOCS method, AOCS Ak4-95. The instrument was calibrated daily using a sealed tube of standard reference canola seed as instructed by the instrument manual. A standard curve of weighed amounts of peanut oil spiked into an inert matrix was run weekly. The variability on repeated analyses was less than or equal to 0.01%.
For fatty acid profile determination, oil was expressed from the samples using a hydraulic press (Fred Carver and Assoc., Wabash, IN). After saponification to release the fatty acids from the triglycerides, the fatty acids in the oil were methylated using a boron-trifluoride catalyst (Bannon et al., 1982). The resulting methyl esters were analyzed using gas chromatography with flame ionization detection (PE Autosampler XL, Perkin Elmer Instruments, Shelton, CN) and quantified as the percent of each fatty acid present of all the fatty acids identified according to the AOCS official method (2004) # Ce 1f-96. Authentic fatty acid methyl esters (Kel Fir FAME 5 mixture, Matreya, LLC, State College, PA) were used for identification by retention times.
Data received were analyzed using R (R Core Team, 2013). Normality of the data was tested using the Shapiro-Wilk test and data that did not follow a normal distribution was subjected to BoxCox analysis to determine appropriate transformations to bring the traits to approximate normality (Box and Cox, 1964). Partitioning of variance was done using mixed model linear regression with the LmerTest package (Kuznetsova et al., 2017) with FDR used for multiple hypothesis correction. Trait summaries were derived from the obtained coefficients. For statistically significant traits (P < 0.05), lines that were significantly different from Fleur 11 (P < 0.05) were determined by running a Dunnett’s multi-comparison test (Dunnett, 1955). The relative effects of introgression on the traits were calculated by taking the difference between the coefficients of each line and Fleur 11 and getting the percentage relative to Fleur 11. PCA biplot analysis was done to examine correlations between the traits that exhibited significant differences among the lines.
Results and Discussion
The spontaneous tetraploidization event that originated cultivated peanut also introduced a reproductive barrier that prevented genetic exchanges with wild diploid relatives. The resultant bottleneck reduced genetic diversity of cultivated varieties. The tetraploid route to interspecific hybridization recently has played a vital role in bridging the ploidy gap and accessing useful alleles from the wild (Simpson, 2001; Fávero et al., 2006). CSSLs, which were developed by the judicious selection of small chromosome segments from the wild in a cultivated background provide an important resource for characterizing the effects of wild alleles on phenotype (Fonceka et al., 2012). In this study, we examined whether wild alleles have an impact on seed quality attributes of a subset of the CSSL population comprising 77 lines that represented approximately 78% of the wild peanut genome. These attributes included 17 fatty acid profiles (palmitoleic, palmitic, margaric, heptadecenoic, stearic, oleic, linoleic, g-linoleic, arachidic, eicosenoic, eicosadienoic, behenic, erucic, lignoceric, cerotic and other acids as well as iodine value), seven sugar profiles (fructose, glucose, myo-inositol, raffinose, stachyose, sucrose and total sugars), O/L ratio, total oil, and protein.
The composition profile of peanut is important as its various physico-chemical constituents contribute to the nutritional, taste, flavor and textural quality of the legume and its products (Ahmed and Ali, 1986; Dwivedi et al., 2000). These traits have a genetic basis, but they also vary depending on environmental factors such as year and area of cultivation as well as the interactions of genetics and environment (Branch et al., 1990; Ku et al., 1998; Andersen and Gorbet, 2002). In line with this, Isleib et al. (2008) in a study of seed composition traits of various breeding lines noted that genetic variation was appreciably high for fatty acid traits in contrast to oil content and sugar profiles, which varied largely as a result of environment. This is consistent with our observation of lower genetic variance for sugar traits compared to the oil components (Table 1).
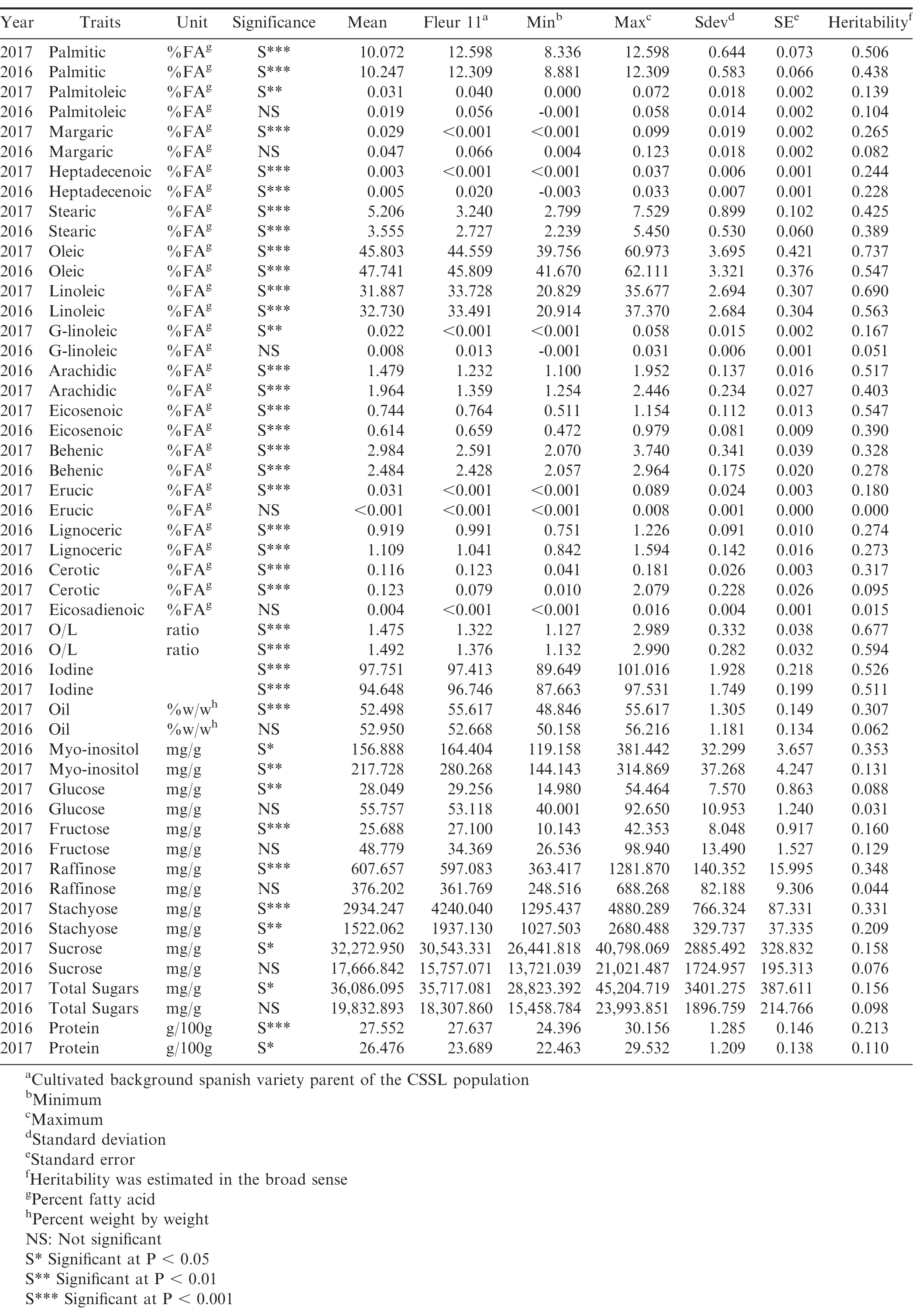
Statistical summary of the chromosome segment substitution lines’ seed composition traits. The lines were developed by introgressing chromosome segments of a synthetic allotetraploid into the background of a cultivated spanish variety of peanut (Fleur 11).
For instance, all but two sugar traits (myo-inositol and stachyose) had no significant genetic differences in 2016 in contrast to 2017 where genetic differences were observed for all traits. This lack of stability across years shows that these sugar profile differences are a product of interactions between genetic and environmental factors rather than just the effects of introgressions. In terms of quantity, sucrose was the most abundant sugar while fructose was least abundant in both years (Table 1). Sucrose contributes to the sweetness attribute of both roasted and unroasted peanut. Upon roasting, reducing sugars are liberated from sucrose, which interact with various amino acids to produce the characteristic nutty flavor associated with roasted peanut (Newell et al., 1967; Mason et al., 1969; Pattee et al., 2000b).
Comparison of the sugars with Fleur 11 identified two lines (CSSL 32 and CSSL 53) that had increased fructose in 2016. CSSL 84 had increased myo-inositol in 2016, CSSL 111 and CSSL 84 had increased raffinose in 2017 and 14 other lines had reduced stachyose in 2017 (Supplementary Table 1). These results do not replicate across the years and are likely due to interaction of the introgressions with environmental factors. In an ideal peanut, the desirable sugar profile would be characterized by high sucrose because of its flavor enhancing properties and reduced raffinose and stachyose (Bishi et al., 2015). Raffinose and stachyose belong to the raffinose family of oligosaccharides (RFOs) which are undesirable since they are not digested by humans due to lack of the α-GAL enzyme. Instead, they are fermented in the large intestines by bacteria resulting in production of hydrogen, carbon dioxide and methane leading to discomforts associated with gassiness (Bryant et al., 2003; Tahir et al., 2012). These sugars are inconsequential for oil production, but significant in food use (Bishi et al., 2013). It would be beneficial to identify lines with genetic variation that increase sucrose and reduce the RFOs.
The mean total oil and protein contents of the CSSLs was 52 and 27% respectively. These values are within the normal range for cultivated peanut (Savage and Keenan, 1994; Grosso and Guzman, 1995; Young and Tai, 2010). While there was overall significant difference among the lines for oil in 2017 and protein in both years (Table 1), none of the individual lines had statistically significant difference from Fleur 11 for both traits. This shows that the introgressions in this population had minimum impact on oil and protein content.
Palmitic, oleic and linoleic acids make up 90% of all fatty acids in peanut. Of these, palmitic acid takes up 10% of the proportion while the unsaturated oleic and linoleic acids make up the remaining 80%. Stearic, lignoceric, behenic, arachidic and eicosenoic acid comprise between 0.02-4.0% of total fatty acids with the rest of the proportion accounted for by other fatty acids (Andersen and Gorbet, 2002). Generally, unsaturated fatty acids can be oxidized resulting in aldehydes, ketones, acids and hydrocarbons that are responsible for diminishing the shelf life and nutritional quality of peanut (Moore and Knauft, 1989; Andersen and Gorbet, 2002). The degree of fatty acid unsaturation results in the classification of the fatty acids as monounsaturated or polyunsaturated, denoting the presence of one or multiple double bonds in the fatty acid side chain respectively. Monounsaturated fatty acids exhibit less oxidation than polyunsaturated fatty acids and are more preferred. Exponential increase in oxidative stability with increasing O/L has been demonstrated (O’Keefe et al., 1993; Davis et al., 2016). Hence, the ratio of the monounsaturated omega 9 oleic acid to the polyunsaturated linoleic acid is a critical quality parameter in peanut. In addition to the quality enhancing property of the trait, many health benefits have also been attributed to this trait including hypotensive and anti-inflammatory effects as well as reduction of type II diabetes and obesity (Terés et al., 2008; Vassiliou et al., 2009; Carrillo et al., 2012).
Oleic, linoleic, palmitic, stearic, arachidic and behenic acid are known to have a strong genetic component (Isleib et al., 2008). The mean values observed in this study (46.7% for oleic, 32.3% for linoleic, 10.2% for palmitic, 4.4% for stearic, 1.7% for arachidic and 2.7% for behenic) (Table 1), were within ranges observed in other studies (Worthington et al., 1972). Oleic acid was inversely related to linoleic, palmitic, behenic, arachidic and stearic acid, with a positive correlation observed with eicosenoic acid. The association of oleic acid with iodine value (a measure of degree of oil unsaturation and hence a measure of oil stability) was inverse (Figure 1). These relationships observed were in general agreement with those of other studies (Mozingo et al., 1989; Hashim et al., 1993; Andersen et al., 1998). Overall, oleic, linoleic and palmitic acid as well as, O/L were the most heritable traits with the heritabilities similar in both years (Table 1). Of the 210 line by trait differences consistent in both years (Table 2), the most represented traits in this comparison were reduction of palmitic acid, increase in arachidic acid and increase in stearic acid with 68, 16 and 5 comparisons, respectively. The lines with the highest number of trait comparisons were CSSL 84, CSSL 100 and CSSL 111, each having 6 comparisons (Table 2). In both years, these lines exhibited reduction in palmitic acid, linoleic acid, and increase in oleic acid, eicosenoic acid and O/L.
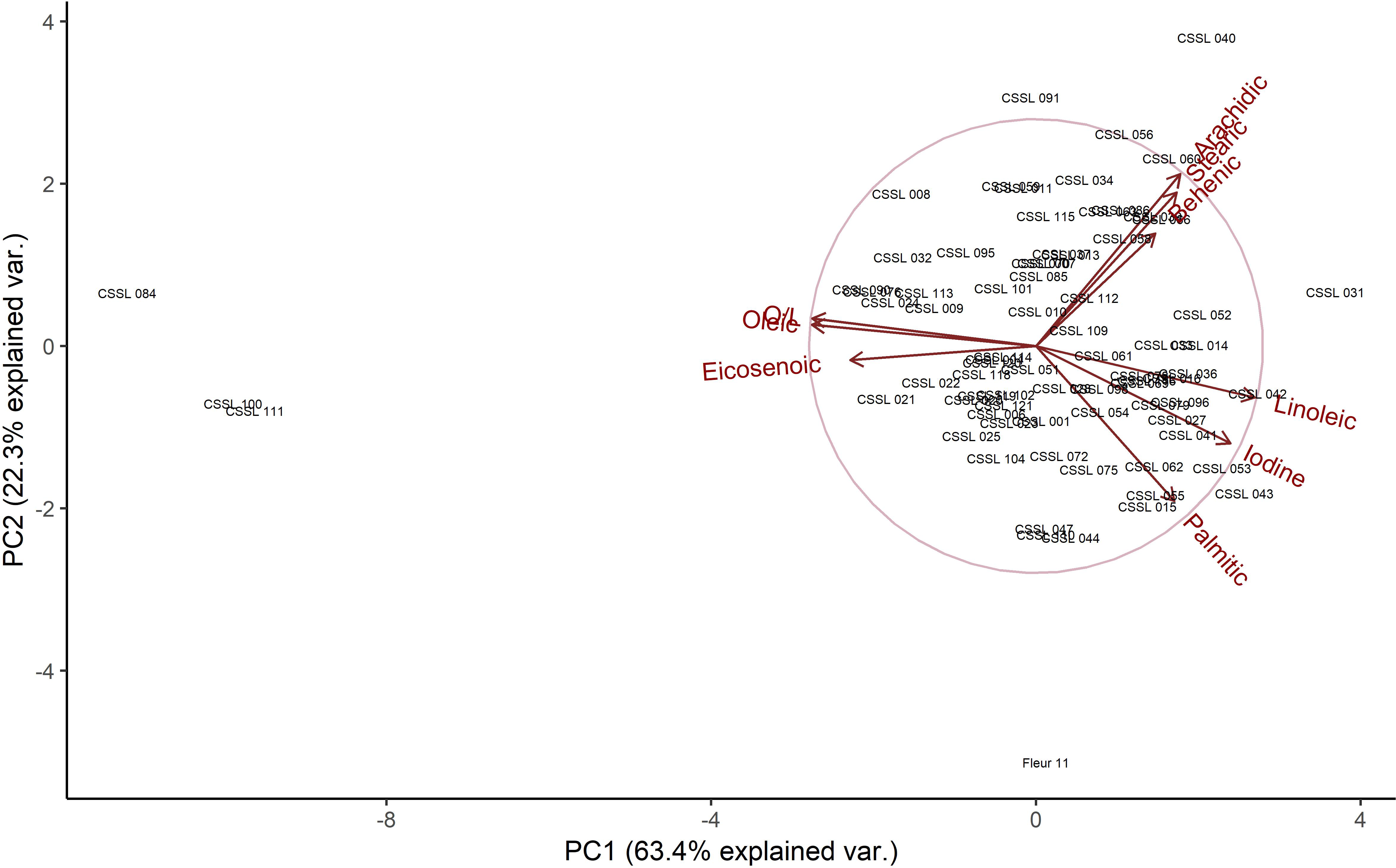
PCA biplot showing the relationships between fatty acids that had significant differences among the chromosome segment substitution lines (CSSLs) in 2016 and 2017. Oleic acid (and hence the O/L) is shown to have an inverse relationship with linoleic, palmitic, behenic, arachidic and stearic acid as well as iodine value. It is positively related to eicosenoic acid. Also observed is a clear separation of CSSL 84, CSSL 100 and CSSL 111 from the rest of the population.
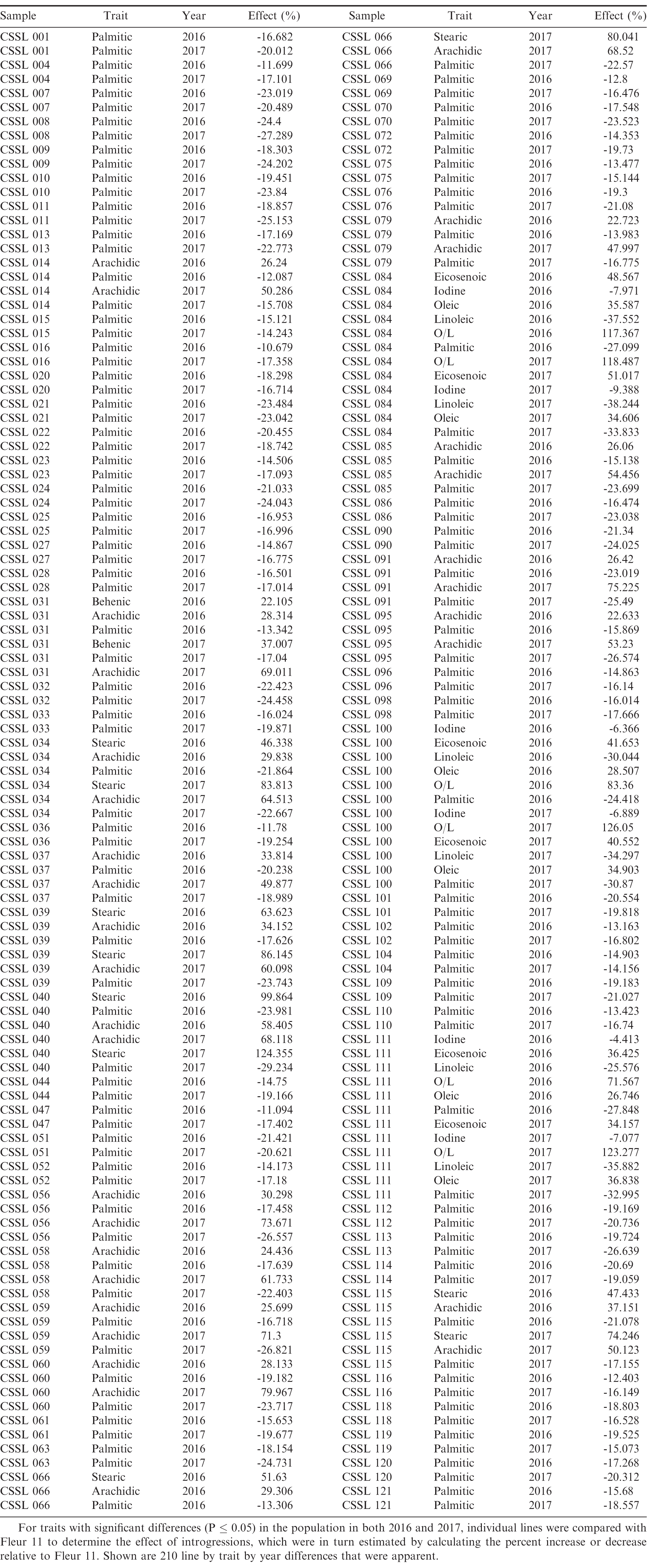
Effects of introgressions relative to Fleur 11 for traits with consistent significant difference in both 2016 and 2017.
The O/L depends on individual concentrations of oleic and linoleic acid. However, the effect of O/L on other fatty acids such as palmitic, total C18 and total saturated fatty acids has also been observed, suggesting a pleiotropic effect. This is in addition to epistatic relationships of the same fatty acids in defining the O/L profile (Isleib et al., 2006). The O/L of Fleur 11 which is the genetic background scaffolding the CSSLs was 1.3. This puts it in the lower spectrum of the normal range of 1.0-2.5 for most commercial cultivated varieties and particularly the spanish types (López et al., 2000; Davis et al., 2016). The mean of the CSSLs was 1.5, indicating that a majority of the lines were normal and similar to Fleur 11 (Table 1). However, three lines, CSSL 84, CSSL 100 and CSSL 111 had ratios of 2.94, 2.76 and 2.66 respectively (Table 3). These values are higher than for the low O/L check New Mexico Valencia A which had a score of 1.21. They are also lower than the score of the high O/L check OLin which had a value of 7.04. Though within the normal range, the values of these higher O/L CSSLs translate to more than 100% increase relative to Fleur 11 which is the standard of comparison (Table 2). The values were also comparable to higher ranked normal O/L runner varieties (Branch et al., 1990; Andersen et al., 1998).
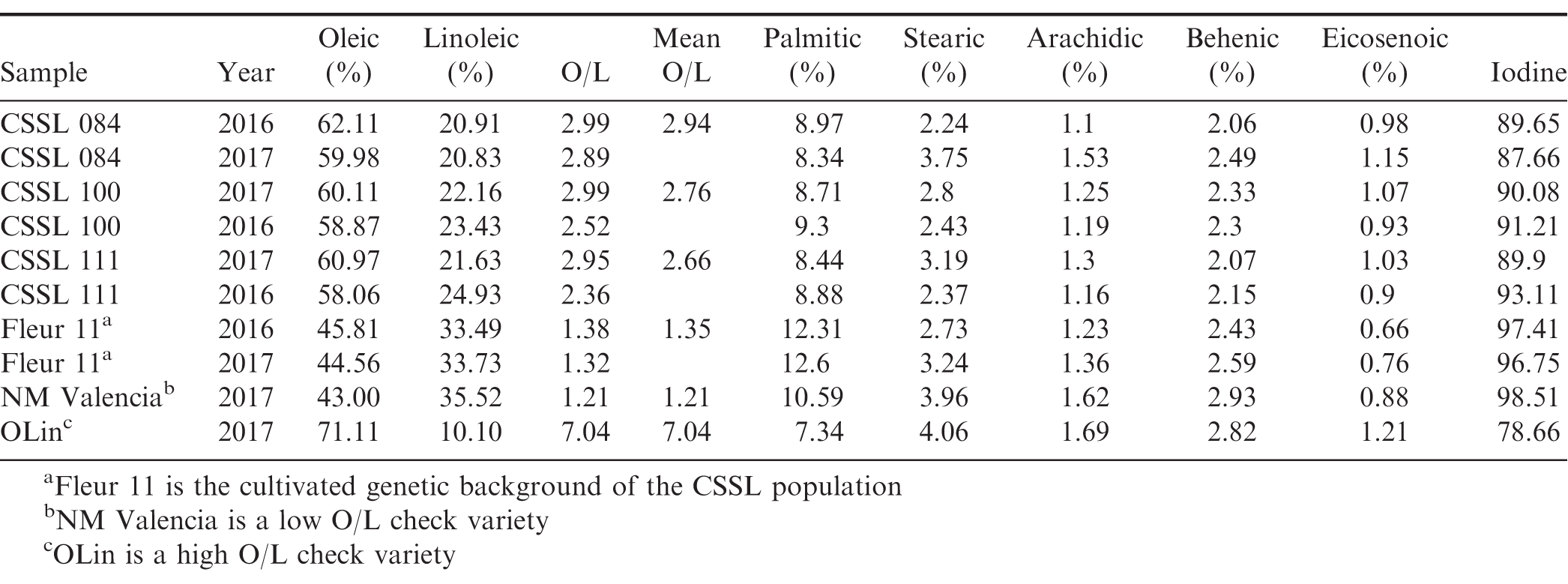
Values of oleic/linoleic and associated fatty acids for the three top lines, CSSL 84, CSSL 100 and CSSL 111.
As noted above, these fatty acid profiles are under genetic control, therefore it follows that introgressions present therein have introduced variations that have increased the O/L. The high O/L trait is the result of two recessive genes ol1 and ol2 also referred to as ahFAD2A and ahFAD2B, respectively (Moore and Knauft, 1989; Jung et al., 2000a; Chu et al., 2009). These genes encode for Δ12-desaturase enzyme that catalyzes the initial step in polyunsaturated fatty acid biosynthesis, which is conversion of oleate to linoleate. In US high O/L cultivars, the mutation in ahFAD2A involves a single base G to A substitution at position 448 (Chu et al., 2007). For the ahFAD2B, either of two mutations achieve desaturase suppression. The first is a position 441_442 A insertion while the second is a MITE insertion at position 665 (Chu et al., 2009). These mutations result in increase of oleic acid above the normal range of 36-67% and reduction of linoleic acid below the normal range of 15-43% (Norden et al., 1987; Moore and Knauft, 1989; Ray et al., 1993). It is likely that introgressions in these three lines may not be associated with the canonical mutations on ahFAD2A and ahFAD2B (Pandey et al., 2014; Shasidhar et al., 2017). Genotyping work on the lines (Gimode et al., unpubl. data) indicates that CSSL 84 has multiple introgressions on nearly all chromosomes, making it difficult to decipher specific polymorphisms that contribute to the trait. CSSL 100 has clear introgressions on the upper and lower arms of chromosomes A10 and B10. There is co-occurrence of polymorphism in these chromosomes between CSSL 100 and CSSL 84. This makes these regions good candidates for further study of the genetic basis for the trait. Unfortunately, clear introgression patterns are yet to be observed in CSSL 111. Presence of multiple introgressions in these lines indicates the likelihood of multiple genes interacting with the genetic background of Fleur 11 to influence variation in the O/L trait as similarly observed by Isleib et al., (2006). The results of this study suggest the potential involvement of new genetic polymorphisms associated with the O/L trait. While these polymorphisms do not radically increase the O/L, understanding their basis may facilitate potentiation of the conventional high O/L trait.
Acknowledgements
The authors would like to thank Jason Golden, Shannon Atkinson, Betty Tyler and Kathy Marchant for technical assistance. The project was funded by USAID Feed the Future Innovation Lab for Peanut. The authors declare no competing interests.
Literature Cited
Ahmed, E. M., & Ali, T. 1986. Textural quality of peanut butter as influenced by peanut seed and oil contents. Peanut Science, 13(1): 18– 20.
Akram, N. A., Shafiq, F., & Ashraf, M. 2018. Peanut (Arachis hypogaea L.): a prospective legume crop to offer multiple health benefits under changing climate. Comprehensive Reviews in Food Science and Food Safety, 17(5): 1325– 1338.
Andersen, P. C., & Gorbet, D. W. 2002. Influence of year and planting date on fatty acid chemistry of high oleic acid and normal peanut genotypes. Journal of Agricultural and Food Chemistry, 50(5): 1298– 1305.
Andersen, P. C., Hill, K., Gorbet, D. W., & Brodbeck, B. V. 1998. Fatty acid and amino acid profiles of selected peanut cultivars and breeding lines. Journal of Food Composition and Analysis, 11(2): 100– 111
AOCS. 2004. Official Methods and Recommended Practices of the American Oil Chemists Society (5th ed.;Firestone, D.Ed.). Champaign, IL: American Oil Chemists Society.
Bailey, K. . 1963. Malnutrition in New Guinean children and its treatment with solid peanut foods. J Journal of Tropical Pediatrics, 9: 35– 43.
Bannon, C. D., Craske, J. D., Hai, N. T., Harper, N. L., & O’Rourke, K. L. 1982. Analysis of fatty acid methyl esters with high accuracy and reliability. II. Methylation of fats and oils with boron trifluoride-methanol. Journal of Chromatography A, 247(1): 63– 69.
Birthal, S. P. ., Rao, P. ., Nigam, S. ., Bantilan, C. ., & Bhagavatulu, S. 2010. Groundnut and Soybean Economies in Asia: Facts, Trends and Outlook. International Crops Research Institute for the Semi-Arid Tropics, Patancheru 502 324, Andhra Pradesh, India. ISBN: 978-92-9066-531-1, p 92.
Bishi, S. K., Kumar, L., Dagla, M. C., Mahatma, M. K., Rathnakumar, A. L., Lalwani, H. B., & Misra, J. B. 2013. Characterization of Spanish peanut germplasm (Arachis hypogaea L.) for sugar profiling and oil quality. Industrial Crops and Products, 51: 46– 50.
Bishi, S. K., Lokesh, K., Mahatma, M. K., Khatediya, N., Chauhan, S. M., & Misra, J. B. 2015. Quality traits of Indian peanut cultivars and their utility as nutritional and functional food. Food Chemistry, 167(15): 107– 114.
Box, G. E. P., & Cox, D. R. 1964. An analysis of transformations. Journal of the Royal Statistical Society: Series B (Methodological), 26(2): 211– 243.
Branch, W. D., Nakayama, T., & Chinnan, M. S. 1990. Fatty acid variation among U.S. Runner-type peanut cultivars. Journal of the American Oil Chemists’ Society, 67(9): 591– 593.
Bryant, R. J., Rao, D. R., & Ogutu, S. 2003. α and β-galactosidase activities and oligosaccharide content in peanuts. Plant Foods for Human Nutrition, 58: 213– 223.
Carrillo, C., Cavia, M. del M. & Alonso-Torre, S. 2012. Role of oleic acid in immune system; mechanism of action; a review. Nutrición Hospitalaria, 27(4): 978– 990.
Chu, Y., Ramos, L., Holbrook, C. C., & Ozias-Akins, P. 2007. Frequency of a loss-of-function mutation in oleoyl-PC desaturase (ahFAD2A) in the mini-core of the U.S. peanut germplasm collection. Crop Science, 47(6): 2372– 2378.
Chu, Y., Wu, C. L., Holbrook, C. C., Tillman, B. L., Person, G., & Ozias-Akins, P. 2011. Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut. The Plant Genome Journal, 4(2): 110.
Chu, Ye Holbrook, C. C., & Ozias-Akins, P. 2009. Two alleles of ahFAD2B control the high oleic acid trait in cultivated peanut. Crop Science, 49(6): 2029– 2036.
Davis, J. P., Price, K. M., Dean, L. L., Sweigart, D. S., Cottonaro, J. M., & Sanders, T. H. 2016. Peanut oil stability and physical properties across a range of industrially relevant oleic acid/linoleic acid ratios. Peanut Science, 43(1): 1– 11.
Dawson, T. P., Perryman, A. H., & Osborne, T. M. 2016. Modelling impacts of climate change on global food security. Climatic Change, 134(3): 429– 440.
Dean, L. L., Hendrix, K. W., Holbrook, C. C., & Sanders, T. H. 2009. Content of Some Nutrients in the Core of the Core of the Peanut Germplasm Collection. Peanut Science, 36(2): 104– 120.
Dunnett, C. W. 1955. A multiple comparison procedure for comparing several treatments with a control. Journal of the American Statistical Association, 50(272): 1096– 1121.
Dwivedi, S. L., Nigam, S. N., & Rao, R. C. N. 2000. Photoperiod effects on seed quality traits in peanut. Crop Science, 40(5): 1223.
Enserink, M. 2008. The peanut butter debate. Science, 322(5898): 36– 38.
FAO., IFAD., UNICEF., WFP., & WHO. 2019. The State of Food Security and Nutrition in the World 2019. Safeguarding Against Economic Slowdowns and Downturns. Rome.Retrieved August 17, 2019, from http://www.fao.org/3/ca5162en/ca5162en.pdf
FAOSTAT. 2014. Retrieved August 17, 2019, from http://www.fao.org/faostat/en/#data/QD/visualize
FAOSTAT. 2017. Retrieved August 17, 2019, from http://www.fao.org/faostat/en/#data/QC
Fávero, A. P., Simpson, C. E., Valls, J. F. M., & Vello, N. A. 2006. Study of the evolution of cultivated peanut through crossability studies among Arachis ipaensis, A. duranensis, and A. hypogaea. Crop Science, 46(4): 1546– 1552.
Fonceka, D., Tossim, H.-A., Rivallan, R., Vignes, H., Lacut, E., de Bellis, F., … Rami, J-F. 2012. Construction of chromosome segment substitution lines in peanut (Arachis hypogaea L.) using a wild synthetic and QTL mapping for plant morphology. PloS One, 7(11): e48642.
Francisco, M. L. D. L., & Resurreccion, A. V. A. 2008. Functional components in peanuts. Critical Reviews in Food Science and Nutrition, 48(8): 715– 746.
Godfray, H. C. J., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., … Toulmin, C. 2010. Food security: The challenge of feeding 9 billion people. Science, 327, 812– 818.
Grosso, N. R., & Guzman, C. A. 1995. Chemical composition of aboriginal peanut (Arachis hypogaea L.) seeds from Peru. Journal of Agricultural and Food Chemistry, 43: 102– 105.
Hashim, I. B., Koehler, P. E., Eitenmiller, R. R., & Kvien, C. K. 1993. Fatty acid composition and tocopherol content of drought stressed florunner peanuts. Peanut Science, 20(1): 21– 24.
Hsi, D. C. H., & Finkner, R. E. 1972. Registration of New Mexico Valencia A peanut (Reg. No. 14). Crop Science, 12(2): 256.
Isleib, T. G., Tillman, B. L., Pattee, H. E., Sanders, T. H., Hendrix, K. W., & Dean, L. O. 2008. Genotype-by-Environment interactions for seed composition traits of breeding lines in the uniform peanut performance test. Peanut Science, 35(2): 130– 138.
Isleib, T. G., Wilson, R. F., & Novitzky, W. P. 2006. Partial dominance, pleiotropism, and epistasis in the inheritance of the high-oleate trait in peanut. Crop Science, 46(3): 1331– 1335.
Israëls, T., Borgstein, E., Jamali, M., De Kraker, J., Caron, H. N., & Molyneux, E. M. 2009. Acute malnutrition is common in Malawian patients with a Wilms tumour: A role for peanut butter. Pediatric Blood and Cancer, 53(7): 1221– 1226.
Janila, P., Manohar, S. S., Patne, N., Variath, M. T., & Nigam, S. N. 2016 a. Genotype × Environment interactions for oil content in peanut and stable high-oil-yielding sources. Crop Science, 56(5): 2506– 2515.
Janila, Pasupuleti Pandey, M. K., Shasidhar, Y., Variath, M. T., Sriswathi, M., Khera, P., … Varshney, R. K. 2016 b. Molecular breeding for introgression of fatty acid desaturase mutant alleles (ahFAD2A and ahFAD2B) enhances oil quality in high and low oil containing peanut genotypes. Plant Science, 242: 203– 213.
Jung, S., Powell, G., Moore, K., & Abbott, A. 2000 a. The high oleate trait in the cultivated peanut [Arachis hypogae L.]. II. Molecular basis and genetics of the trait. Molecular and General Genetics, 263(5): 806– 811.
Jung, S., Swift, D., Sengoku, E., Patel, M., Teulé, F., Powell, G., … Abbott, A. 2000 b. The high oleate trait in the cultivated peanut [Arachis hypogaea L.]. I. Isolation and characterization of two genes encoding microsomal oleoyl-PC desaturases. Molecular and General Genetics, 263(5): 796– 805.
Ku, K. L., Lee, R. S., Young, C. T., & Chiou, R. Y. Y. 1998. Roasted peanut flavor and related compositional characteristics of peanut kernels of spring and fall crops grown in Taiwan. Journal of Agricultural and Food Chemistry, 46(8): 3220– 3224.
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. 2017. lmerTest package: tests in linear mixed effects models. Journal of Statistical Software, 82(13): 1– 26.
Lobell, D. B., Schlenker, W., & Costa-Roberts, J. 2011. Climate trends and global crop production since 1980. Science, 333(6042): 616– 620.
López, Y., Nadaf, H. L., Smith, O. D., Connell, J. P., Reddy, A. S., & Fritz, A. K. 2000. Isolation and characterization of the Δ12-fatty acid desaturase in peanut (Arachis hypogaea L.) and search for polymorphisms for the high oleate trait in Spanish market-type lines. Theoretical and Applied Genetics, 101(7): 1131– 1138.
Mason, M. E., Newell, J. A., Johnson, B. R., Koehler, P. E., & Waller, G. R. 1969. Nonvolatile flavor components of peanuts, 17:728. Journal of Agricultural and Food Chemistry.
Mehta, D. 2018. The Green Revolution did not increase poverty and hunger for millions. Nature Plants, 4: 736.
Moore, K. M., & Knauft, D. A. 1989. The inheritance of high oleic acid in peanut. Journal of Heredity, 80(3): 252– 253.
Mozingo, R. W., Coffelt, T. A., & Wynne, J. C. 1989. Market grade effects on fatty acid composition of five peanut cultivars. Agronomy Journal, 80(1): 73.
Myer, R. O., Johnson, D. D., Knauft, D. A., Gorbet, D. W., Brendemuhl, J. H., & Walker, W. R. 1992. Effect of feeding high-oleic-acid peanuts to growing-finishing swine on resulting carcass fatty acid profile and on carcass and meat quality characteristics. Journal of Animal Science, 70(12): 3734– 3741.
Nawade, B., Mishra, G. P., Radhakrishnan, T., Dodia, S. M., Ahmad, S., Kumar, A., … Kundu, R. 2018. High oleic peanut breeding: Achievements, perspectives, and prospects. Trends in Food Science and Technology, 78: 107– 119.
Newell, J. A., Mason, M. E., & Matlock, R. S. 1967. Precursors of typical and atypical roasted peanut flavor. Journal of Agricultural and Food Chemistry, 15(5): 767– 772.
Norden, A. J., Gorbet, D. W., Knauft, D. A., & Young, C. T. 1987. Variability in oil quality among peanut genotypes in the Florida breeding program. Peanut Science, 14(1): 7– 11.
O’Byrne, D. J., Knauft, D. A., & Shireman, R. B. 1997. Low fat-monounsaturated rich diets containing high-oleic peanuts improve serum lipoprotein profiles. Lipids, 32(7): 687– 695.
O’Keefe, S. F., Wiley, V. A., & Knauft, D. A. 1993. Comparison of oxidative stability of high- and normal-oleic peanut oils. Journal of the American Oil Chemists’ Society, 70(5): 489– 492.
Pandey, M.K., Wang, M.L., Qiao, L., Feng, S., Khera, P., … Guo, B. 2014. Identification of QTLs associated with oil content and mapping FAD2 genes and their relative contribution to oil quality in peanut (Arachis hypogaea L.). BMC Genetics, 15(1): 1– 14.
Pattee, H. E., Isleib, T. G., Giesbrecht, F. G., & McFeeters, R. F. 2000 a. Investigations into genotypic variations of peanut carbohydrates. Journal of Agricultural and Food Chemistry, 48(3): 750– 756.
Pattee, H. E., Isleib, T. G., Giesbrecht, F. G., & McFeeters, R. F. 2000 b. Relationships of sweet, bitter, and roasted peanut sensory attributes with carbohydrate components in peanuts. Journal of Agricultural and Food Chemistry, 48(3): 757– 763.
Pingali, P. L. 2012. Green revolution: Impacts, limits, and the path ahead. Proceedings of the National Academy of Sciences of the United States of America, 109: 12302– 12308.
R Core Team. 2013. R: A Language and Environment for Statistical Computing.
Ray, T. K., Holly, S. P., Knauft, D. A., Abbott, A. G., & Powell, G. L. 1993. The primary defect in developing seed from the high oleate variety of peanut (Arachis hypogaea L.) is the absence of Δ12-desaturase activity. Plant Science, 91(1): 15– 21.
Savage, G. P., & Keenan, J. I. 1994. The composition and nutritive value of groundnut kernels. In Smartt J.(eds.) The Groundnut Crop. World Crop Series. Springer, Dordrecht.
Shasidhar, Y., Vishwakarma, M.K., Pandey, M.K., Janila, P., Variath, M.T., … Varshney, R. K. 2017. Molecular mapping of oil content and fatty acids using dense genetic maps in groundnut (Arachis hypogaea L.). Frontiers in Plant Science, 8: 794.
Simpson, C. E. 2001. Use of Wild Arachis species/introgression of genes into A. hypogaea L. Peanut Science, 28(2): 114– 116.
Simpson, C. E., Baring, M. R., Schubert, A. M., Melouk, H. A., Lopez, Y., & Kirby, J. S. 2003. Registration of ‘OLin’ peanut. Crop Science, 43(5): 1880– a.
Tahir, M., Båga, M., Vandenberg, A., & Chibbar, R. N. 2012. An assessment of raffinose family oligosaccharides and sucrose concentration in genus lens. Crop Science, 52(4): 1713– 1720.
Terés, S., Barceló-Coblijn, G., Benet, M., Alvarez, R., Bressani, R., Halver, J. E., & Escribá, P. V. 2008. Oleic acid content is responsible for the reduction in blood pressure induced by olive oil. Proceedings of the National Academy of Sciences of the United States of America, 105(37): 13811– 13816.
Toomer, O. T. 2018. Nutritional chemistry of the peanut (Arachis hypogaea). Critical Reviews in Food Science and Nutrition, 58: 3042– 3053.
Variath, M. T., & Janila, P. 2017. Economic and academic importance of peanut. In Varshney, R.,Pandey, M., & Puppala, N.(eds.) The Peanut Genome. Compendium of Plant Genomes (pp. 7–26).
Vassiliou, E. K., Gonzalez, A., Garcia, C., Tadros, J. H., Chakraborty, G., & Toney, J. H. 2009. Oleic acid and peanut oil high in oleic acid reverse the inhibitory effect of insulin production of the inflammatory cytokine TNF-alpha both in vitro and in vivo systems. Lipids in Health and Disease, 8: 25.
Worthington, R. E., Hammons, R. O., & Allison, J. R. 1972. Varietal differences and seasonal effects on fatty acid composition and stability of oil from 82 peanut genotypes. Journal of Agricultural and Food Chemistry, 20(3): 727– 730.
Young, C. T., & Tai, Y. P. 2010. Variations in percent protein of individual seeds from five peanut plants. Agronomy Journal, 66(3): 439– 440.
Notes
- First and sixth authors: Former Graduate Student and Professor, Institute of Plant Breeding Genetics and Genomics, University of Georgia, Tifton, GA 31793; Second author: Research Professional, Department of Horticulture, University of Georgia, Tifton, GA 31793; Third author: Food Technologist, Market Quality and Handling Research Unit, USDA, North Carolina State University, Raleigh NC 27695; Fourth author: Supervisory Research Geneticist, United States Department of Agriculture - Agricultural Research Service, Tifton GA, 31793; Fifth author: Researcher and Scientific Coordinator, CERAAS, Thies, Senegal. [^] *Corresponding author Email: pozias@uga.edu
Author Affiliations