

Introduction
Peanut (Arachis hypogaea L.) is an important oilseed crop grown worldwide. This crop originated in South America and is grown in mostly tropical, subtropical and relatively warm temperate conditions (Hammons et al., 2016). Variable environmental conditions encountered from the time of planting until harvest, from the broad regions peanuts are cultivated, can greatly influence stand establishment, crop development, and yield. Air and soil temperatures as well as soil moisture affect germination and early seedling growth. Considering adequate soil moisture at planting and in the early season along with proper management practices, air and soil temperatures play an important role in stand establishment and seedling development. Thus, selection of planting date is a critical factor for production. Peanut seed emergence occurs between 6 to 11 d after planting, depending upon soil and air temperatures (Canavar and Kaynak, 2010). As a general rule, peanut germination is considered optimum in the soil temperature range of 20 to 35 C at a 10 cm soil depth for three consecutive d (Kvien et al., 2019) and air temperatures between 27 and 32 C are considered optimum for peanut growth and yield (Boote et al., 1989; Williams and Boote, 1995).
Gardner and Auma (1989) reported a negative effect of delaying peanut planting on plant growth such as plant height and leaf area. Similarly, Prasad et al. (2006) evaluated the effect of different planting dates with soil temperatures ranging from 15 to 32 C on emergence and early season growth of six peanut cultivars. The authors reported the lowest emergence rate and smallest seedlings for the planting date with the coolest temperature conditions.
Temperature is the driving force for several chemical reactions occurring in plant cells at different stages throughout the life cycle. Decreases in temperature below the optimum range generally decrease the rate of chemical reactions. Sub-optimal temperature increases the probability of early-season seedling injury from pests and pathogens in peanuts with potential seedling death, ultimately resulting in reduced plant populations and lower yield potential (Prasad et al., 2006; Bell et al., 1993; Bell, 1986). Leong and Ong (1983) reported a linear increase in rate of peanut development with increasing temperature from 19 to 31 C. Low temperature in the early season can also have pronounced effects on the physiology and biochemistry of seedlings, resulting in reduced plant growth and development due to lower photosynthetic rates (Allen and Ort, 2001). Reduced net assimilation rate under low temperature can be due to stomatal factors, such as stomata closure or non-stomatal factors, such as deactivation of Rubisco (Holaday et al. 1992; 2016). Bagnall et al. (1988) reported 50 to 70% decrease in net photosynthesis within the first two d with a change in temperature from 30 to 19 C. The authors further indicated that the decrease in net photosynthesis under sub-optimal temperature conditions was due to non-stomatal limitations.
Supra-optimal temperature at planting and in the early season can also negatively affect germination and seedling growth (Challinor et al., 2005; Hamidou et al., 2013). For peanuts, air temperatures near 30 C are considered optimal for growth and physiological processes such as photosynthesis (Wood, 1968; Cox 1979; Ong, 1984). Current evidence suggests that high temperature limits photosynthesis by inhibiting rubisco activity through deactivation of rubisco activase, by limiting photosynthetic electron transport via disruptions in the thylakoid membrane, or by decreased affinity of rubisco for carbon dioxide and low carbon dioxide to oxygen ratio inside the leaf, thereby increasing photorespiration rate (Feller et al., 1998; Crafts-Brandner and Salvucci, 2000; Salvucci and Crafts- Brander, 2004; Schrader et al., 2004; Wise et al., 2004).
Following emergence, first true leaves are the first green tissues in peanuts that are photosynthetically active, contributing substantially to production of photoassimilates used for further whole plant growth and development. The amount of energy produced by the leaves through photosynthesis depends upon leaf size and photosynthetic efficiency. The net carbon available to drive growth is influenced by photosynthetic pigments available for light energy capture, rate of thylakoid reactions, efficiency of carbon fixation reactions and rate of dark respiration (Taiz and Zeiger, 2010).
The first true leaves develop at the node next to node zero, designated as node one (Boote, 1982). During the early stage of crop growth, leaf area index of peanut seedlings is low (Taiz and Zeiger, 2010). However, as growth progresses, plants develop more leaves and expand existing leaves, increasing the leaf area index. At a later stage, such as the onset of flowering, peanut plants have a substantial number of leaves above the first true leaves. Variation in photosynthetic rate in different leaves within a peanut plant was reported, with higher photosynthetic rates in young, fully expanded leaves than older lower-canopy leaves (Henning et al., 1979). However, studies characterizing the contributions of first true leaves to peanut plant growth at different developmental stages or growth temperature conditions are limited. Therefore, the objectives of this study were to evaluate the contribution of first true leaves of modern peanut cultivars to whole-plant growth under different temperature conditions during the early season and at the onset of flowering as well as to identify the photosynthetic components more closely associated with photosynthetic efficiency of the first true leaves. It was hypothesized that first true leaf development will have a pronounced effect on peanut plant growth under supra-optimal temperature conditions, irrespective of plant developmental stage. In addition, non-stomatal components associated with the thylakoid reactions will more closely drive photosynthetic efficiency of first true leaves in modern peanut cultivars than stomatal factors.
Materials and Methods
Plant material and experimental layout
A two-year field experiment was conducted at the Lang-Rigdon Research Farm of the University of Georgia, Tifton, GA (N 31°52', W 83°55', 108 masl) during the growing seasons in 2017 and 2018. The soil series at the site is Tifton loamy sand (fine-loamy, kaolinitic, thermic, Plinthic Kandiudults). Three runner-type peanut cultivars, Georgia-06G (Branch, 2007), Georgia-14N (Branch and Brenneman, 2015) and TifNV-High O/L (Holbrook et al., 2017), were planted on three different dates corresponding to April (early), May (optimal) and June (late) in order to generate differences in temperature conditions at planting and in the early season. For the 2017 growing season, the selected planting dates were April 17, May 10, and June 5 and for the 2018 season, the planting dates were April 25, May 14, and June 11. A split-plot design with planting date as the whole-plot and cultivar as the sub-plot was used, and treatments were replicated four times. For both growing seasons, plots were four single-rows spaced 91 cm apart and 11 m long. Seeds were sown at a depth of approximately 5 cm with a seeding rate of 19 seeds per linear meter. Soil and air temperatures as well as precipitation data were collected from the University of Georgia weather station closest to the experimental location. Fungicide applications followed the schedule for a high-risk category to help protect the trial from damaging level from disease incidence (Kemerait et al., 2018). Overhead sprinkler irrigation was used to irrigate the field according to the University of Georgia Extension recommendations (Porter, 2019). Plant growth and physiological parameters were collected at two sampling dates, three and five weeks after planting (WAP), for each planting date.
Measurements and sampling protocol
Seedling emergence index
Emergence was recorded daily during the first 14 d after planting (DAP) from a 3 m section of the two middle rows in each plot. The total number of emerged seedlings were counted, with seedlings being considered emerged only when visible first true leaves were completely above the soil surface. The seedling emergence index (SEI) was calculated according to equation described in Maguire (1962).
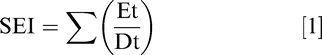
Whole plant growth
Whole plants (not including tissues below the cotyledons) from a total of 1.8 m were destructively sampled from the two middle rows of each plot between 0700 and 0800 h at 3 and 5 WAP. Plastic bags with moist paper tower were used to store the harvested plants in order to avoid wilting. The samples were kept in a refrigerator at 4 C until further processing. Measurements included number of plants per 1.8-m section, first true leaf area (FTLA), and tissue dry weight. The first true leaves were separated from the rest of the leaves and leaf area was recorded using a table-top leaf area meter (LI-3100; LI-COR, Lincoln, NE). Then, all the leaves (including the first true leaves) and stems were dried at 80 C for 48 h using a forced air oven and dry weight (DW) of leaves and stems per plant (in g/plant) was recorded.
Physiological Measurements
Pigment content
Pigment contents (chlorophyll A, chlorophyll B, and total carotenoids) were quantified in first true leaf samples at 3 and 5 WAP. Four 5-mm diameter leaf discs were placed in amber vials filled with 5 ml reagent grade ethanol solution (96% ethanol solution) and incubated at 4 C for fourteen d. Afterwards, sample absorbance was read at 665, 649, and 470 nm wavelengths using a multi-well plate reader (Synergy HTX, BioTek, Winooski, VT). Content of chlorophyll A, chlorophyll B, and total carotenoids per unit leaf area were calculated using the equations described in Lichtenthaler and Wellburn (1983) and expressed in μg/cm2.
Gas exchange and fluorescence measurements
Along with first true leaf pigment content, gas exchange and fluorescence were recorded at 3 and 5 WAP on single leaflets per leaf (top leaflets further from peduncle insertion). Light-adapted measurements of gas exchange, chlorophyll fluorescence, and leaf temperatures were determined in the first true leaves between 1100 and 1400 h using the LI-6400XT portable photosynthesis system (LI-COR, Nebraska, USA) and the fluorometer chamber (Model LI-6400-40, LI-COR, Lincoln, NE). The gas exchange measurements included net photosynthesis (An), intercellular CO2 concentration (Ci), stomatal conductance (gs), and leaf temperature (C). The reference CO2 concentration in the leaf chamber was set as 400 μmol/mol along with photosynthetically active radiation (PAR) of 1500 μmol m/s. Other chamber settings included a flow rate of 500 μmol/s, block temperature matching the ambient air temperature at the time of measurements and relative humidity of 60 ± 10%. The 6400-40 Leaf Chamber Fluorometer was used to record chlorophyll fluorescence measurements. After the steady state net photosynthesis (An) and fluorescence (Fs) were reached, the multi-phase protocol was used to determine the maximum fluorescence intensity (Fm'), at which all reaction centers are closed (Earl and Ennahli, 2004). Further, as per the equations given in Maxwell and Johnson (2000) and Genty et al. (1989), actual quantum yield of photosystem II (ΦPSII) and electron transport rate through photosystem II (ETR) were calculated as:
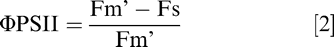
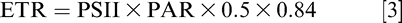
Dark-adapted measurements, including midday dark respiration (Rd), were obtained in the first true leaves by covering some plants from each plot with a structure consisting of a frame and opaque black felt. The plants were dark-adapted for approximately 2 h from 0930 to 1130 h prior to the measurements. For the dark-adapted measurements, all chamber settings were the same as those used for the light-adapted measurements, except that no illumination was provided in the dark measurements. Gross photosynthesis (Ag) was then calculated by summing Rd (obtained from the dark-adapted assessment) and An (obtained from the light-adapted assessment). Gross photosynthesis was expressed in μmol m/s.
From the measured gas exchange and fluorescence parameters, further parameters were derived according to the methods described in Ennahli and Earl (2005), Pilon et al. (2018), and Galmés et al. (2007). The derived parameters are the electron transport rate to carbon assimilation (ETRa; in μmol m/s), electron transport rate to photorespiration or rate of oxygenation reaction of rubisco (ETRp; μmol m/s), carbon dioxide concentration in chloroplast (Cc; μmol/mol), and mesophyll conductance (gm; mol m/s).
Statistical Analysis
A two-way analysis of variance was conducted using JMP Pro 13.0 (SAS Institute, 2016). Planting date and cultivar were considered fixed effects and data from the two years were analysed separately as differences were observed between the years. Replication and replication × planting date were considered as random effects. Means were separated using Fisher's Least Significant Difference post hoc test at the 0.05 probability level whenever significant differences were observed. Pairwise correlations were also performed to identify associations between plant growth and the physiological component processes.
Results and Discussion
Uniform and vigorous seedling emergence ensures a successful start to the peanut crop. Along with seed characteristics, environmental conditions influence the rate and time of seedling emergence. Under optimum soil moisture, soil temperature plays a significant role in seedling emergence. For both the growing seasons, April planting dates corresponded to the lowest temperature conditions, whereas June planting dates had the highest temperatures conditions. In 2017, average 10-cm soil temperature within the first 14 d after planting ranged from 21 to 27 C for April, 24 to 30 C for May, and 24 to 30 C for June. In 2018, temperatures were similar to 2017 in April, but they were higher for May and June, with average 10-cm soil temperatures between 22 and 28 C for April, 25 and 27 C for May, and 28 an 33 C for June.
Seedling emergence index was unaffected by planting date by cultivar interaction, but was significantly impacted by planting date (Fig. 1; P = 0.01) and cultivar (Fig. 2; P = 0.01) separately in both growing seasons. In 2017, May conditions promoted higher initial seedling emergence rate, followed by April and June (Fig. 1A). In 2018, higher emergence index was observed in May and June compared to April (Fig. 1B). In addition, GA-06G resulted in the highest seedling emergence index across all cultivars in both growing seasons (Fig. 2), which is the cultivar with higher seed weight (Tubbs, 2019). Studies on other species indicated that emergence and development of seedlings are influenced by seed size (Snider et al., 2014; Ries and Everson, 1973). The lower emergence index observed for GA-14N in 2017 can be attributed to the lower seed vigor of this specific cultivar (which has smaller seeds; Tubbs, 2019) due to adverse conditions encountered during 2016 growing season. Seedlings that emerge early have a greater chance of survival and higher relative yield as compared to seedlings that emerge later (Wanjura et al., 1969). Soil temperatures between 27 and 33 C, encountered by the seeds planted in May and June, promoted higher emergence rate of the peanut cultivars.
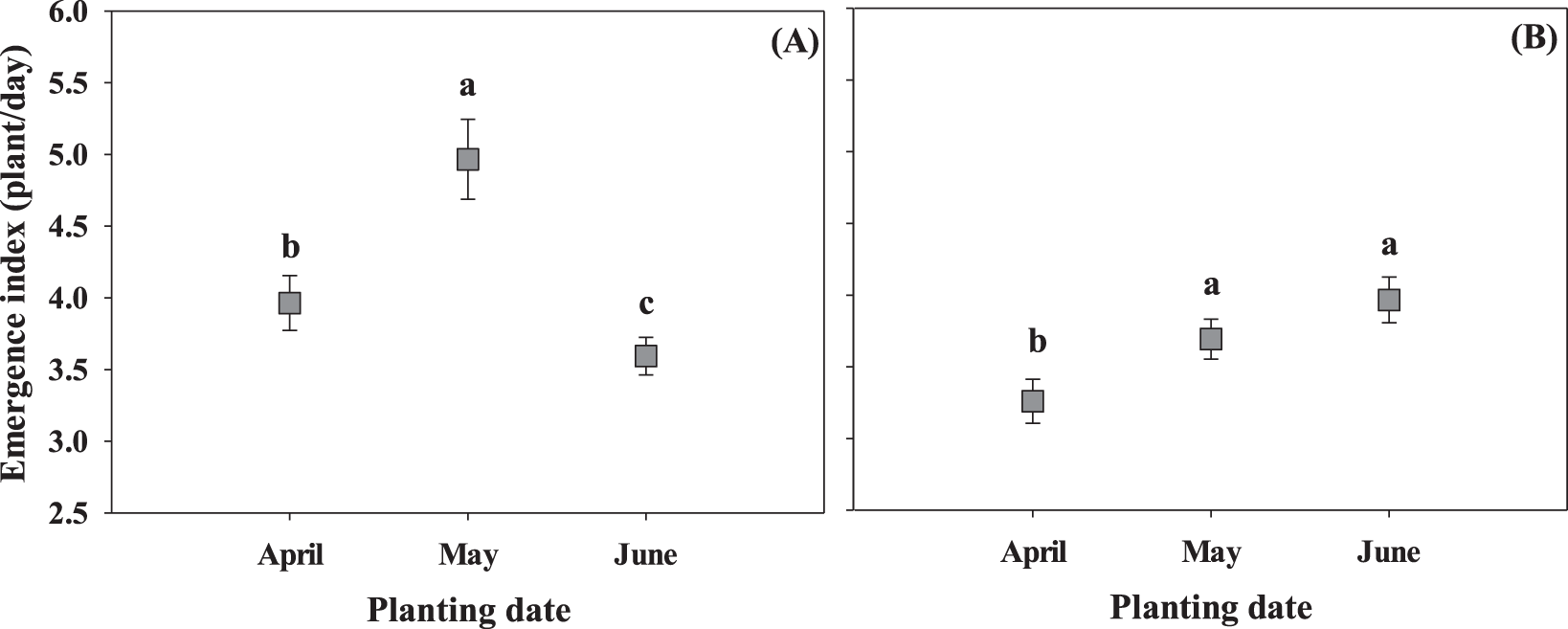
Seedling emergence index (SEI) for three planting dates, April, May, and June, at 3 and 5 weeks after planting in two seasons, 2017 (A) and 2018 (B). Values represent means ± standard error with data combined across three cultivars within planting dates (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
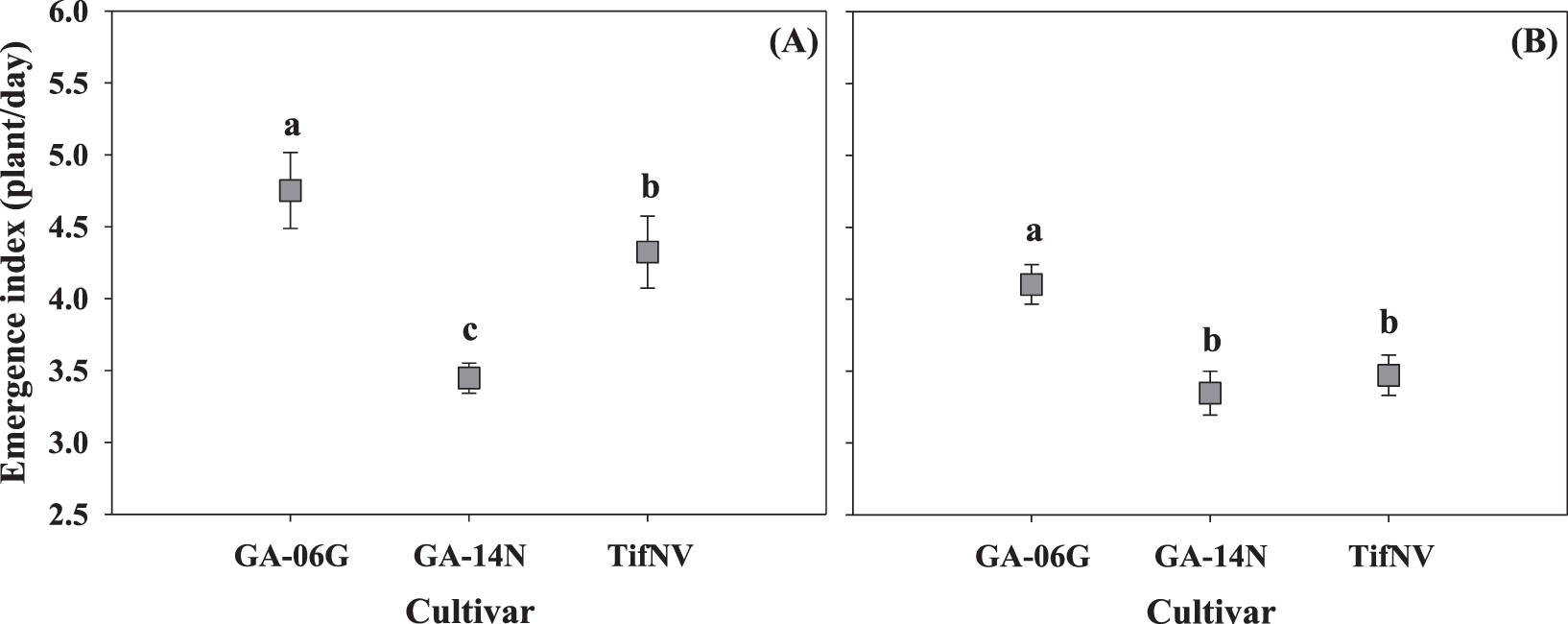
Seedling emergence index (SEI) for peanut cultivars, GA-06G, GA-14N, and TifNV, at 3 and 5 weeks after planting in two seasons, 2017 (A) and 2018 (B). Values represent means ± standard error with data combined across three planting dates within cultivar (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
First true leaf area and whole-plant dry weight were not affected by planting date by cultivar interaction at either 3 or 5 WAP for both growing seasons. However, a significant effect of planting date was observed for the measured growth parameters at both sampling times in 2017 and 2018, except for the first true leaf area at 3 WAP in 2017 (Fig. 3 and Table 1). First true leaf area was greater in June than May and April at all times (Fig. 3). For instance, in 2018, FTLA in June was, on average, 15% higher than May and April at 3 WAP, whereas at 5 WAP, June planting resulted in 36% and 30% greater FTLA than April and May, respectively (Fig. 3B). Total plant dry weight was affected by planting dates at 3 and 5 WAP in both growing seasons (Table 1). Temperature conditions for the June planting resulted in the highest dry matter accumulation, followed by May and lastly April, with the lowest dry matter accumulation. At 5 WAP in 2017, total dry weight for the June planting date was approximately 2.9-fold higher than April and 1.4-fold higher than May (Table 1). Similarly, in 2018, June resulted in 2.7-fold higher total dry weight per plant compared to April and May. Other research reported similar effect of low temperature conditions on peanut growth and development (Leong and Ong, 1983; Allen and Ort, 2001). Leong and Ong (1983) suggested a linear increase in rate of growth and development of peanuts with increasing temperature from 19 to 31 C. Jordan et al. (2019) reported an increase in population and growth of peanut plants with planting shift from mid-April to end of May.
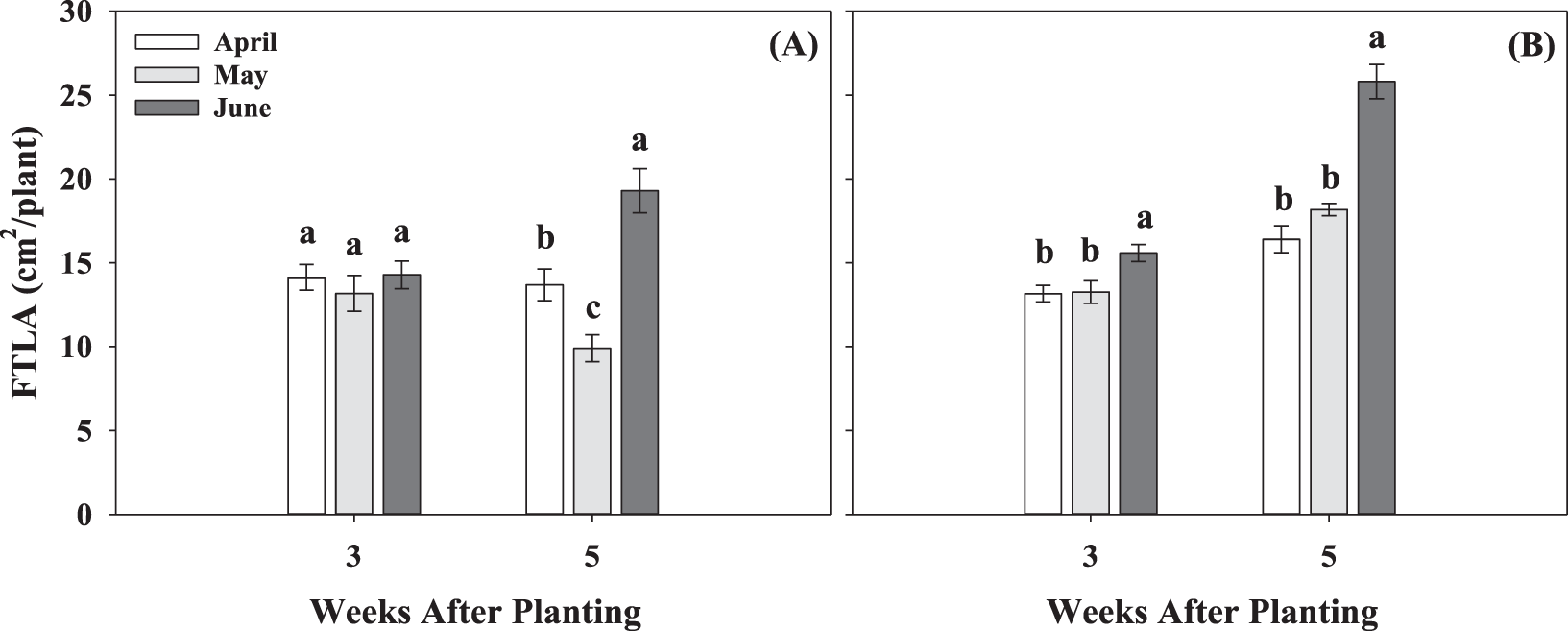
First true leaf area per plant (FTLA; cm2/plant) for three planting dates, April, May, and June, at 3 and 5 weeks after planting in two seasons, 2017 (A) and 2018 (B). Values represent means ± standard error with data combined across three cultivars within planting dates (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
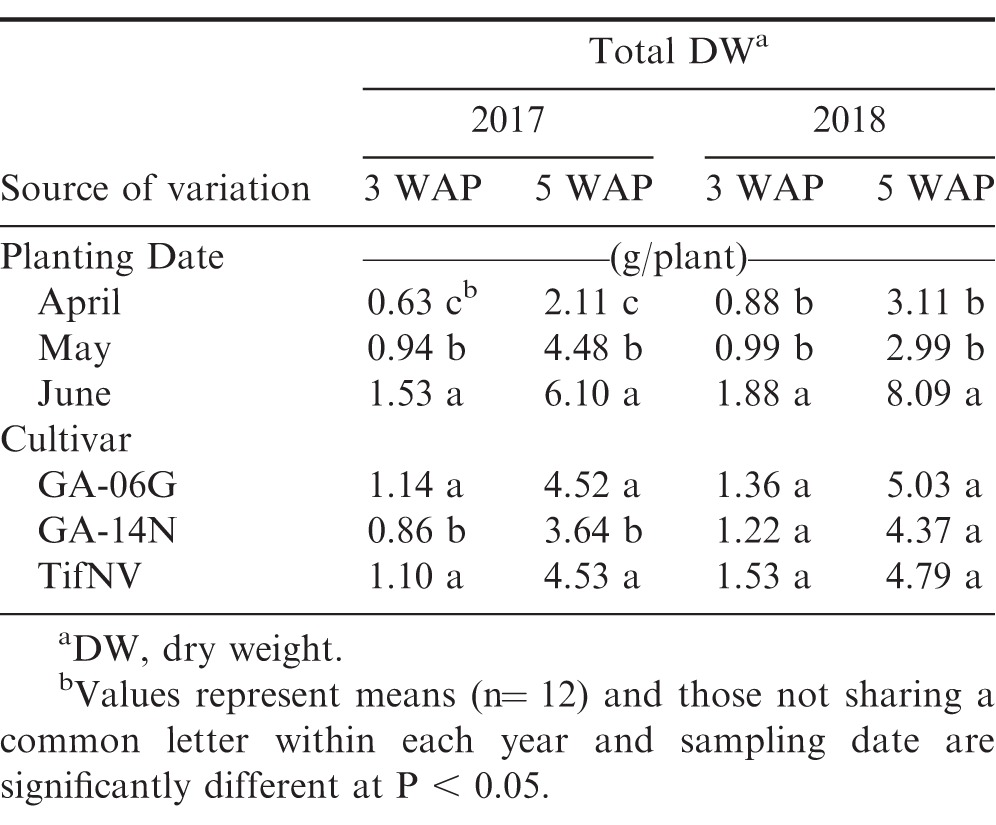
Dry weight measurements at 3 and 5 weeks after planting (WAP) for the three selected planting dates, April, May, and June, and for three peanut cultivars, Georgia-06G, Georgia-14N, and TifNV-High O/L, during two growing seasons, 2017 and 2018.
First true leaf area was also significantly affected by cultivar at 3 and 5 WAP in 2017 (Fig. 4). TifNV had the highest FTLA at both sampling times, followed by GA-06G and lastly GA-14N. More specifically, FTLA of TifNV was, on average, 25% and 34% greater than GA-06G and GA-14N, respectively (Fig. 4). Prasad et al. (2006) studied the effect of different soil temperatures on seedling emergence and early season growth of six peanut cultivars. The authors reported lower germination rates and early season growth for planting dates corresponding to coolest sowing temperatures. Also, differences in early vigor among the cultivars were due to differences in rate of first true leaf differentiation, which resulted in significant differences in early seedling development among different peanut cultivars.
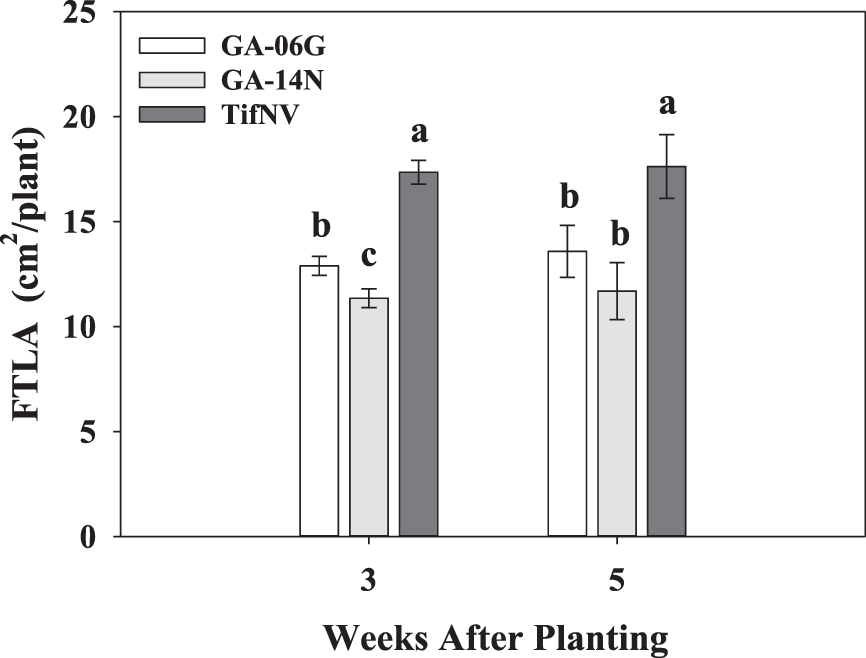
First true leaf area per plant (FTLA/plant) for peanut cultivars, GA-06G, GA-14N, and TifNV, at 3 and 5 weeks after planting in 2017. Values represent means ± standard error with data combined across three planting dates within cultivar (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
The current research indicates that temperature conditions substantially impacted overall peanut plant growth, including first true leaf development (Table 1 and Fig. 3). Average daily temperature calculated from each planting date to 3 WAP (2017 and 2018 averaged) varied from 13.7 to 26.7 C, 19.5 to 29.9 C, and 21.4 to 31 C for April, May, and June, respectively. Moreover, the average daily temperatures from planting to 5 WAP were 15.5 to 28.2 C for April, 19.7 to 29.9 C for May, and 21.7 to 31.6 C for June. Rapid development and differentiation of first true leaves is an indication of greater seedling vigor in other crops, such as cotton (Gossypium hirsutum L.; Pilon et al., 2016). Dry weight is also a commonly used parameter to indicate plant vigor (Snider et al., 2016). Pairwise correlations showed positive relationship between first true leaf area and whole-plant dry matter at early season (r = 0.42) and at the onset of flowering (r = 0.52), suggesting contribution of the first true leaf development to peanut plant vigor (Tables 2 and 3).

Pairwise correlation coefficients for growth parameters, pigment contents and physiological parameters at 3 WAP for three cultivars, three planting dates, and two growing seasons combined.
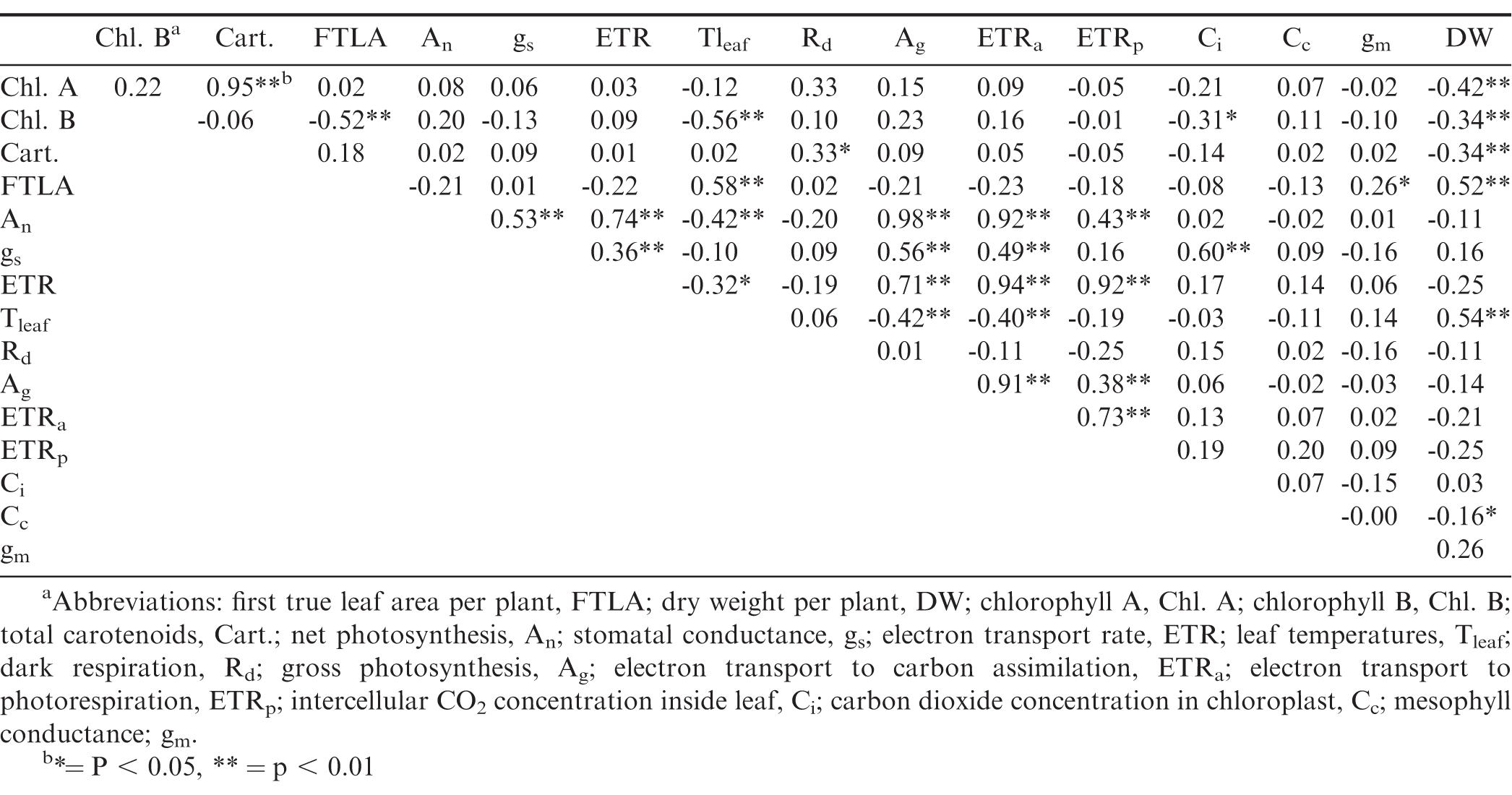
Pairwise correlation coefficients for growth parameters, pigment contents and physiological parameters at 5 WAP for three cultivars, three planting dates, and two growing seasons combined.
In addition to FTLA, pigment content (chlorophyll A and total carotenoids), Tleaf, gs, and Ci measured on the first true leaf were also positively correlated with whole-plant dry weight during early growth stage (Table 2). However, whole-plant dry weight was negatively correlated with multiple photosynthetic variables measured on the first true leaf such as net photosynthesis, ETR, ETRa, and ETRp. Net photosynthesis along with ETR decreased with an increase in plant dry weight. In addition, there was a positive correlation between plant dry matter and Rd of first true leaves. At the onset of flowering, Tleaf (r = 0.354) was positively correlated with whole-plant dry weight, whereas negative relationship was observed between whole-plant dry weight and measured pigment concentrations and Cc (Table 3). No significant correlation was observed between whole-plant dry weight and net photosynthesis at 5 WAP. First true leaf area development is a potential indicator of whole-plant vigor at early season and the onset of flowering; however, the photosynthetic efficiency of the first true leaf does not have direct impact on whole-plant growth, because An was negatively associated with whole-plant dry matter at early stage and not associated at the onset of flowering, when first true leaves were fully developed and the plant had a high number of upper canopy leaves photosynthetically active.
No significant planting date by cultivar interation was observed for any of the pigment contents collected at 3 and 5 WAP in the 2017 and 2018 seasons. However, all pigment contents were significantly affected by planting date at 3 and 5 WAP in both seasons, except for chlorohyll B at 5 WAP in 2018 (Fig. 5). At 3 WAP in the 2017 season, the April planting resulted in the highest chlorophyll A and total carotenoids, followed by May and June plantings (Fig. 5A and E). Chlorophyll B at 3 WAP in 2017 was higher for the May planting date compared to April and June (Fig. 5C). A negative impact of delayed planting was observed at 3 and 5 WAP, where all the measured pigment contents were the lowest for June. For example, at 5 WAP, chlorophyll A was 38% lower in June compared with April (Fig. 5A). Similarly, total carotenoids were 52% lower in June than April planting (Fig. 5E). In 2018, samples for pigment analysis were not collected at 3 WAP for the first planting date (April) due to rainy conditions. June resulted in substantially higher content of all pigments measured when compared to May at 3 WAP (Fig. 5B, D and F). Similar to trends observed in the 2017 season, a negative effect of delayed planting on pigment contents was observed at 5 WAP for the 2018 season. Specifically, chlorophyll A and chlorophyll B were 1.7- and 1.2-fold higher, respectively, for the April planting compared to June (Fig. 5B and D). Synthesis of leaf chlorophyll is known to decrease with increase in temperature, especially above 32 C, which likely accounted for the lower pigment content in plants planted in June compared with those planted in April. In addition, pigment content was affected by cultivar in 2018, except for chlorophyll A and total carotenoids at 3 WAP (Fig. 6). At 3 WAP, TifNV had, on average, 62% higher chlorophyll B than GA-06G and GA-14N (Fig. 6B). At 5 WAP, all measured pigment contents were greater for GA-06G and TifNV compared to GA-14N (Fig. 6).
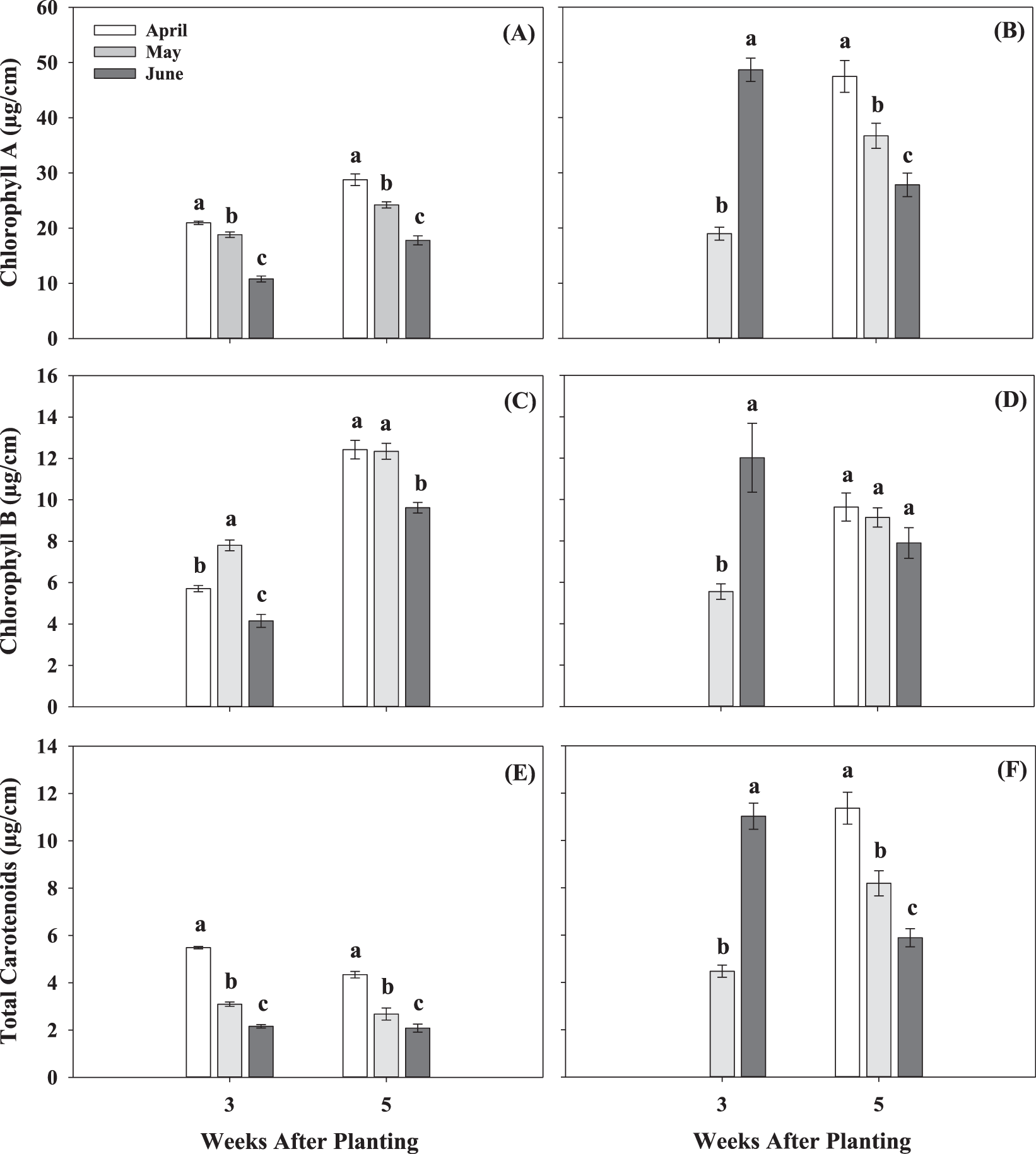
Pigment contents, chlorophyll A, chlorophyll B, and total carotenoids, for the three planting dates, April, May, and June, at 3 and 5 weeks after planting for two growing seasons, 2017 (A, C, E) and 2018 (B, D, F). Values represent means ± standard error with data combined across three cultivar within planting date (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
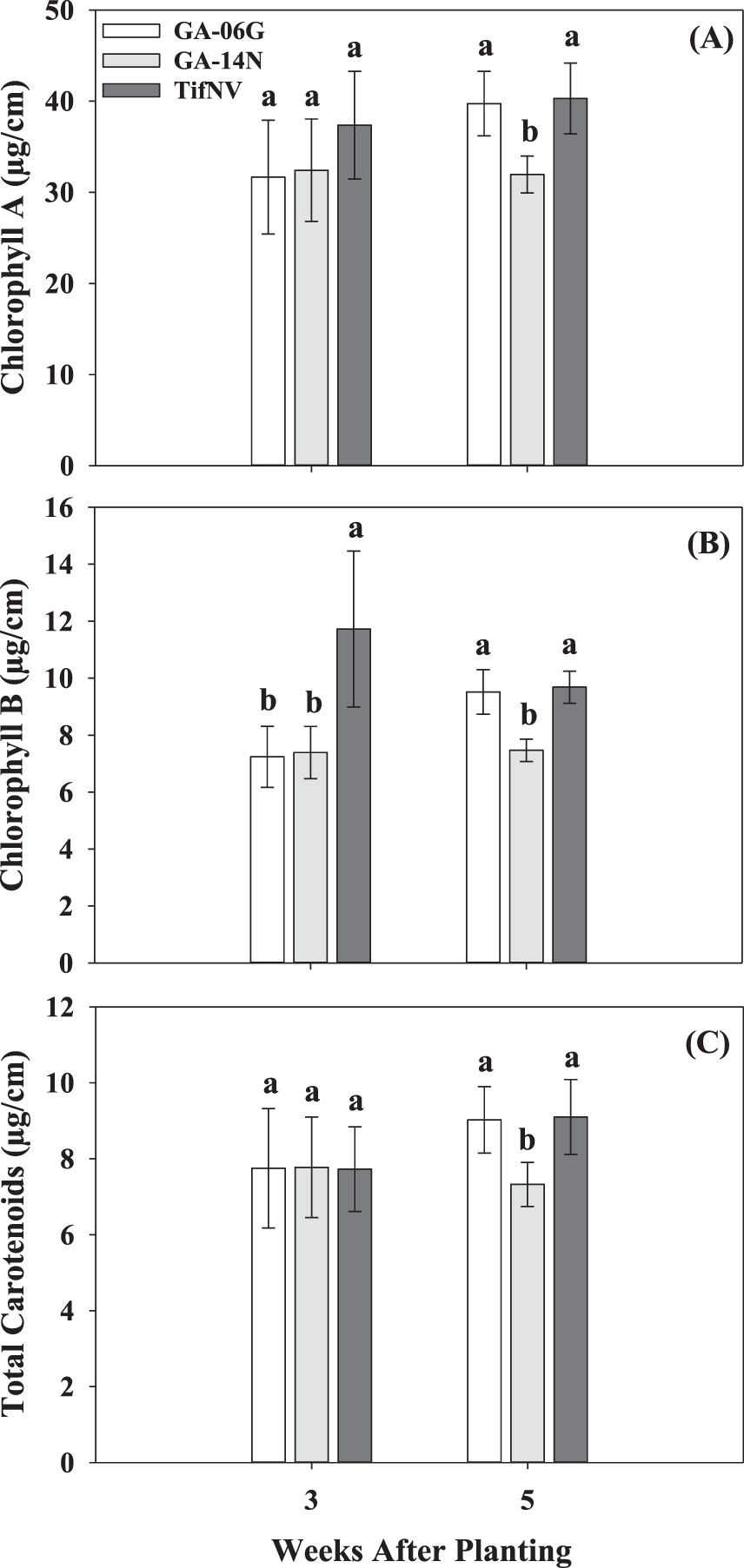
Pigment contents, chlorophyll A, chlorophyll B, and total carotenoids, at 3 and 5 weeks after planting for the three peanut cultivars, Georgia-06G, Georgia-14N, and TifNV-High O/L, for the 2018 growing season. Values represent means ± standard error with data combined across three planting dates within cultivar (n = 12), except for data across 3 WAP, in which n = 8 Values not sharing a common letter within each week are significantly different at P < 0.05.
Leaf temperature was also significantly impacted by different planting dates (Fig. 7). Increases in leaf temperature were observed with delayed planting. For example, at 5 WAP in 2017, leaf temperature for the June planting was, on average, 10 C higher than April (Fig. 7A). In addition, at 5 WAP in 2018, albeit at a lesser extent, the June planting resulted in the highest leaf temperature of 33.1 C compared to average leaf temperature of 30 C in April (Fig. 7B). Extreme temperatures (soil, air, and leaf) can affect photosynthesis, thus affecting growth and development. Ketring (1986) reported a reduction in photosynthesis in peanut leaves with further reduction in growth when leaf temperature was above 35 C. June plantings resulted in the highest temperatures and, although none of the planting conditions promoted leaf temperatures above 35 C, significant negative correlations were observed between leaf temperature and net photosynthesis (Tables 2 and 3). Net photosynthesis decreased with the increase in leaf temperature at 3 WAP (r = -0.46) and 5 WAP (r = -0.42) when the data was combined across both growing seasons. Cox (1979) suggested 28 C as optimum temperature for growth during early development and onset of flowering in peanut plants. Additionally, the authors indicated absence of growth at night temperatures of 15.5 C. The temperatures corresponding to the April planting were closer to optimal temperatures (28 C) for early growth and development of peanuts compared to larger deviation from optimum temperatures for the May and June plantings. Electron transport rate to carbon assimilation of the first true leaf was also negatively correlated with leaf temperature, irrespective of sample date (Tables 2 and 3). Higher temperatures encountered by plants in the June planting resulted in negative impact on photosynthetic efficiency of first true leaves, mainly affecting the thylakoid reactions.
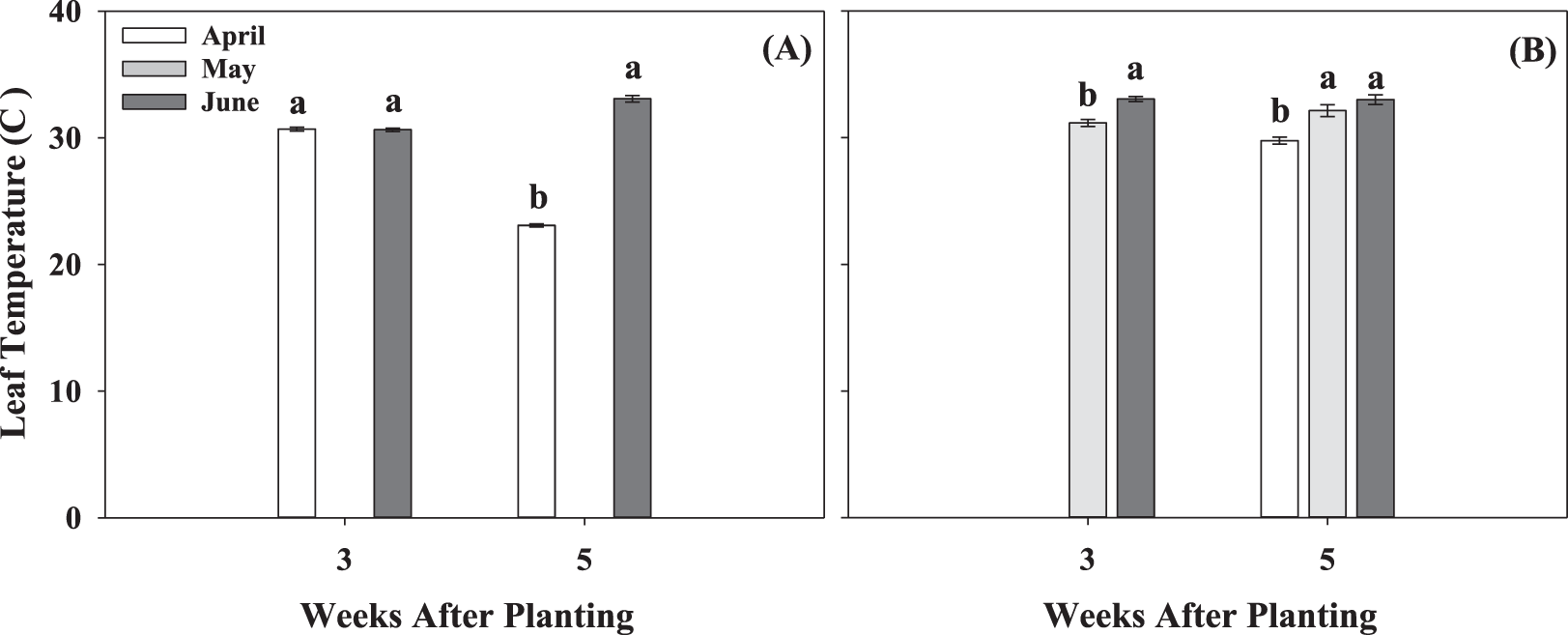
Leaf temperature at 3 and 5 weeks after planting for the three planting dates, April, May, and June, for two growing seasons, 2017 (A) and 2018 (B). Values represent means ± standard error with data combined across three cultivars within planting date (n = 12). Values not sharing a common letter within each week are significantly different at P < 0.05.
Net photosynthesis of the first true leaf was unaffected by planting date, cultivar, and planting date x cultivar interaction at all sampling dates on both growing seasons (Table 4). However, a significant planting date effect was observed for various photosynthetic components, including ETR, ETRa, ETRp, Ci, Cc, and gs for the 2017 season, and ETRp, Ci, and gs for the 2018 season (Tables 4 and 5). At early season, ETR, ETRa, and ETRp were greater for the April planting compared to June planting, whereas Ci, Cc, and gs were higher for June compared to April in 2017 (Tables 4 and 5). In 2018, ETRp, Ci and gs were greater for June than those for May planting date. However, at the onset of flowering, planting date affected only Ci in 2017, with higher values for June planting compared to April, and ETRp in 2018, with higher values for April and May compared to June (Tables 4 and 5). Although photosynthetic components of first true leaves were affected by different temperature conditions promoted by the different planting dates, net photosynthesis remained unaffected. In order to identify which photosynthetic components were more closely associated with photosynthesis maintenance under different environmental conditions, pairwise correlations were obtained among photosynthetic components of the thylakoid reactions as well as the carboxylation reactions on the first true leaves. Positive correlations were observed between net photosynthesis of the first true leaves and multiple photosynthetic components, such as ETR, ETRa, ETRp, and gs (Tables 2 and 3). At early growth, net photosynthesis was strongly associated with Ag (r = 0.98) and ETRa (r = 0.84), while a weaker correlation was observed between An and gs (r = 0.56; Table 2). Similarly, at the onset of flowering, net photosynthesis showed a strong correlation with Ag (r = 0.98) and ETRa (r = 0.92) and was weakly associated with gs (r = 0.52; Table 3). Stomatal and non-stomatal factors affected photosynthetic responses of first true leaves to temperature; however, stronger influence of thylakoid components was observed on net photosynthesis at early season and the onset of flowering, as evidenced by strong, positive correlations with ETRa and weak association with gs as well as no significant relationship with Ci and Cc (Tables 2 and 3).
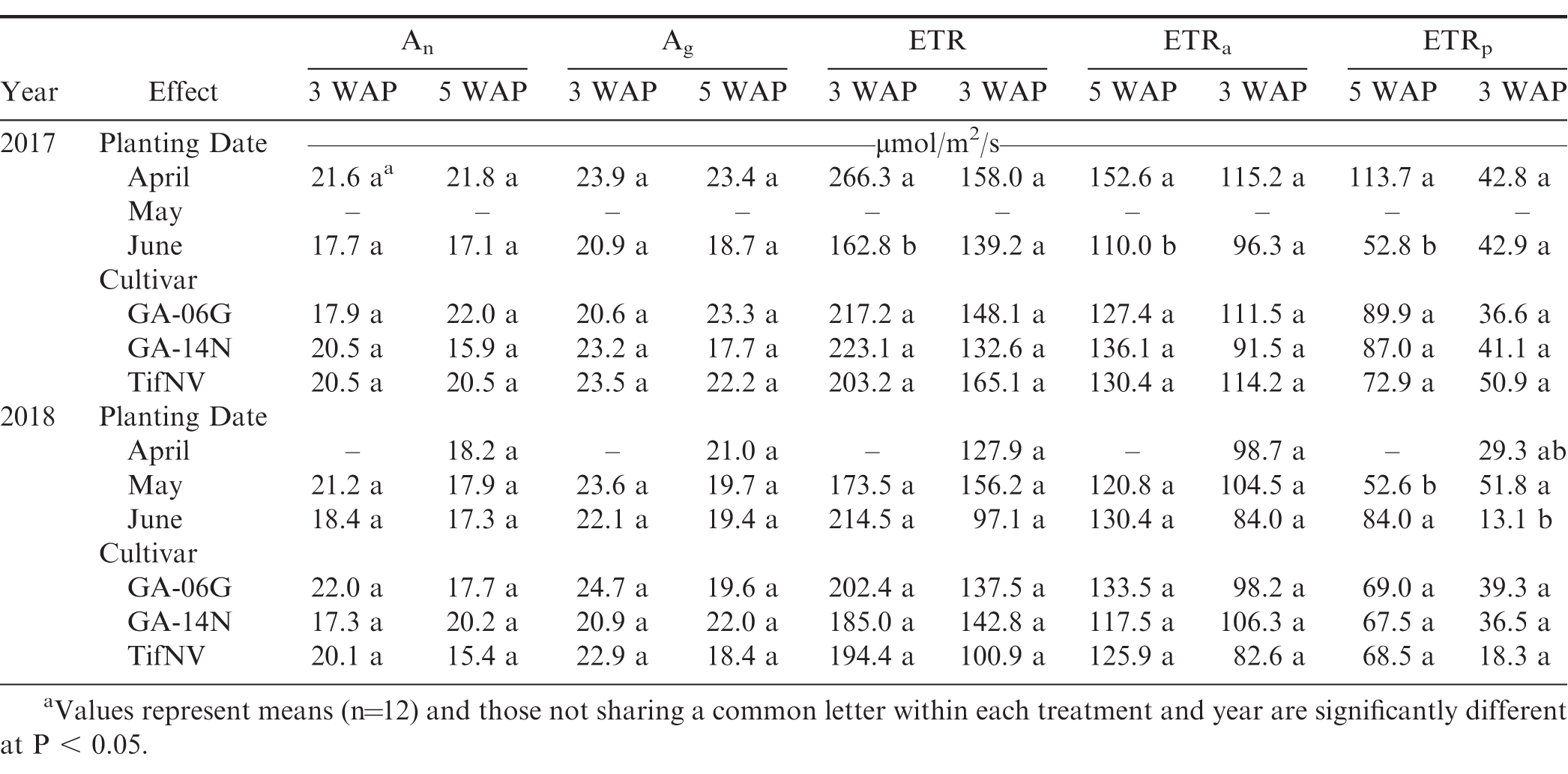
Net photosynthesis (An), gross photosynthesis (Ag), electron transport rate (ETR), electron transport to carbon assimilation (ETRa), and electron transport to photorespiration (ETRp) at 3 and 5 weeks after planting (WAP) for the three planting dates, April, May, and June, and three peanut cultivars, Georgia-06G, Georgia-14N, and TifNV-High O/L, for 2017 and 2018 seasons.
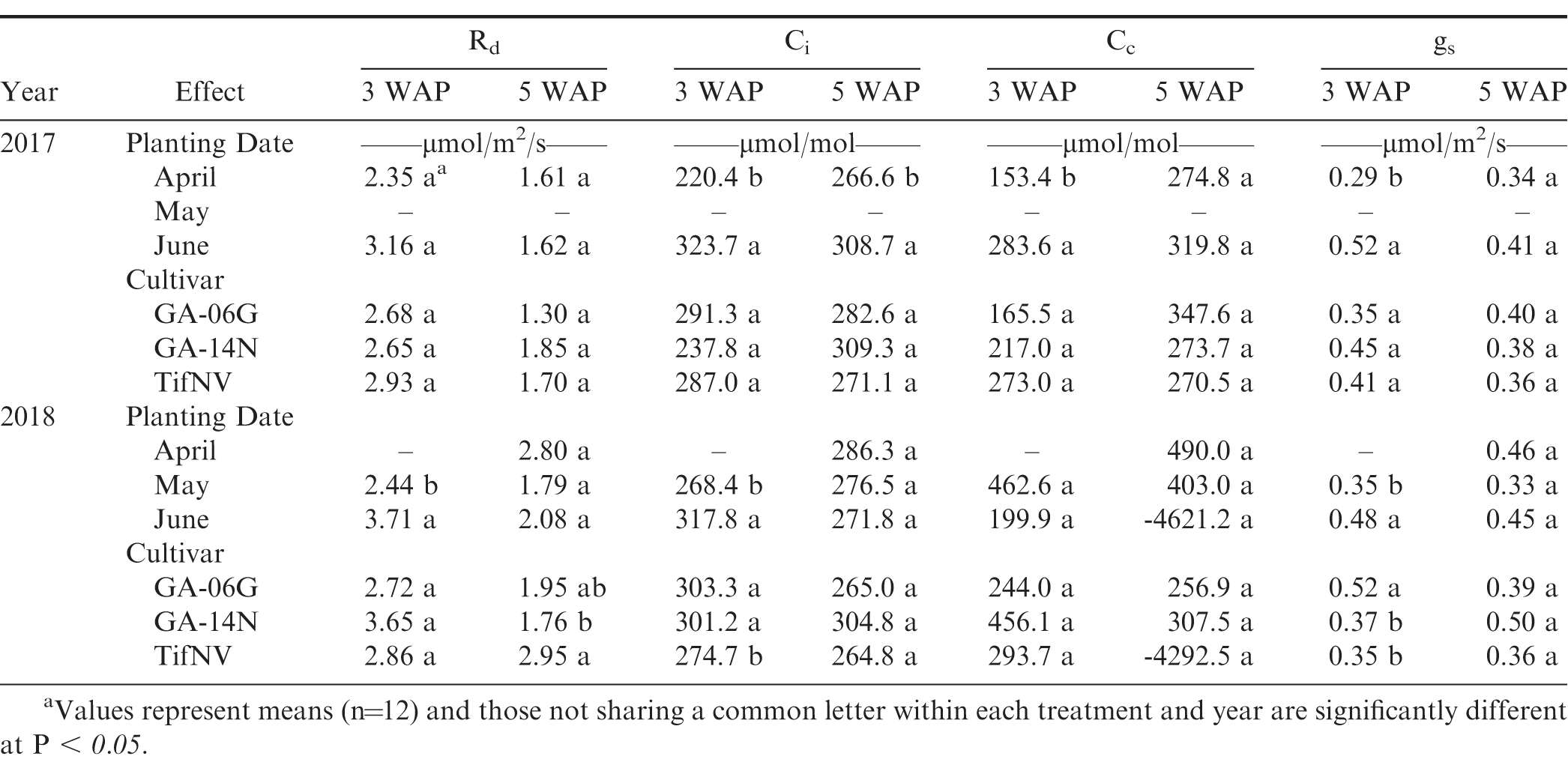
Midday dark respiration (Rd), intercellular CO2 concentration inside leaf (Ci), CO2 concentration in the chloroplast (Cc), stomatal conductance (gs), and mesophyll conductance (gm) at 3 and 5 weeks after planting (WAP) for the three planting dates, April, May, and June, and three cultivars, Georgia-06G, Georgia-14N, and TifNV-High O/L, for both 2017 and 2018 seasons.
Summary and Conclusions
It is concluded that a rapid development of first true leaves of peanut plants contribute to whole-plant growth at early season and at the onset of flowering under adverse temperature conditions due to increased surface area available for light interception. However, the photosynthetic efficiency of the first true leaf has no direct impact on first true leaf surface area, irrespective of the growth stage. Furthermore, net photosynthesis of first true leaves was unaffected by temperature conditions at early season and even at the onset of flowering, when the plants have multiple upper leaves photosynthetically active, mainly due to improved activity of non-stomatal components associated with the thylakoid reactions. Whole-plant growth is more strongly associated with development of first true leaf than photosynthetic efficiency of those leaves in the peanut crop.
Acknowledgements
The authors would like to thank Georgia Peanut Commission for the financial support of the project. The authors also thank our technicians Aaron Moore and Michael Purvis and the peanut team members Mario Scolari, Beatriz Fabreti, and Nuengsap Thangthong for help and technical support.
Literature Cited
D.J Allen, and D.R Ort (2001). Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci 6: 36- 42.
D.J., Bagnall, R.W King, and G.D Farquhar (1988). Temperature-dependent feedback inhibition of photosynthesis in peanuts. Planta 175: 348- 354.
M Bell, (1986). Effect of sowing date on growth and development of irrigated peanuts (Arachis hypogaea L.) cv. Early Bunch, in a monsoonal tropical environment. Aust. J. Agric. Res 37: 361- 373.
M.J., Bell, G.C Wright, and G.R Harch (1993). Environmental and agronomic effects on the growth of four peanut cultivars in a sub-tropical environment. II. Dry matter partitioning. Exp. Agric 29: 491- 501.
K.J Boote, (1982). Growth stages of peanuts (Arachis hypogea L.). Peanut Sci 9: 35- 46.
Boote, K.J., J.W Jones, G Hoogenboom, G.G Wilkerson, and S.S Jagtap 1989 PNUTGRO V1.02: Technical Documentation. IBSNAT Project Department of Agronomy and Soil Science, University of Hawaii. Honolulu.
W.D Branch, (2007). Registration of 'Georgia-06G' Peanut. J. of Plant Registrations 1: 120.
W.D Branch, and T Brenneman (2015). Registration of 'Georgia-14N' Peanut. J. of Plant Registration 9: 159.
A.J., Challinor, T.R Wheeler, P.Q Craufurd, and J.M Slingo (2005). Simulation of the impact of high temperature stress on annual crop yields. Agric. For. Meteorol 135: 180- 189.
F.R Cox, (1979). Effect of temperature treatment on peanut vegetative and fruit growth. Peanut Sci 6: 14- 17.
S.J Crafts-Brandnerand M.E Salvucci (2000). Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc. Natl. Acad. Sci. USA 97: 13430- 13435.
H.J Earl, and S Ennahli (2004). Estimating photosynthetic electron transport via chlorophyll fluorometry without photosystem II light saturation. Photosynth. Res 82: 177- 186.
S Ennahli, and H.J Earl (2005). Physiological limitations to photosynthetic carbon assimilation in cotton under water stress. Crop Sci 45: 2374- 2382.
U., Feller S.J., Crafts-Brandnerand M.E Salvucci (1998). Moderately high temperatures inhibit ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activase-mediated activation of Rubisco. Plant Physiol 116: 539- 546.
J., Galmés, H Medrano, and J Flexas (2007). Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New Phytologist 175: 81- 93.
F.P Gardner, and E.O Auma (1989). Canopy structure, light interception and yield and market quality of peanut genotypes as influenced by planting pattern and planting date. Field Crops Res 20: 13- 29.
B., Genty, J.M Briantais, and N.R Baker (1989). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 990: 87- 92.
F., Hamidou, O Halilou, and V Vadez (2013). Assessment of groundnut under combined heat and drought stress. J. Agron. Crop Sci 199: 1- 11.
Hammons, R.O., D Herman, H.T Stalker 2016 Origin and early history of the peanut Pages: 1- 26 In: Peanuts: Genetics, Processing, and Utilization AOCS Press, San Diego, CA.
R.J., Henning, R.H Brown, and D.A Ashley (1979). Effects of leaf position and plant age on photosynthesis and translocation in peanut I. Apparent photosynthesis and l4C translocation. Peanut Sci 6: 46- 50.
A.S., Holaday, J.R Mahan, and P Payton (2016). Molecular Biology and Physiology. J. Cotton Sci 20: 220- 231.
A.S., Holaday, W Martindale, R Aired, A.L Brooks, and R.C Leegood (1992). Changes in activities of enzymes of carbon metabolism in leaves during exposure of plants to low temperature. Plant Physiol 98: 1105- 1114.
C.C., Holbrook, P. O Akins, Y Chu, A.K Culbreath, C Kvien, and T Brenneman (2017). Registration of 'TifNV-High O/L' Peanut. J. Plant Registration 11: 228- 230.
B.S., Jordan, A.K Culbreath, T.B Brenneman, R.C Kemeraitand K.L Stevenson (2019). Effect of planting date and peanut cultivar on epidemics of late leaf spot in Georgia. Plant Dis 103: 990- 995.
Kemerait, R., A.K Culbreath, E Prostko, T Brenneman, R.S Tubbs, R Srinivasan, M Abney, W.S Monfort, A Rabinowitz, B Tillman, N Dufault, D Rowland, M Mulvaney, I Small, A Hagan, J Sarver, D Anco, and N Smith 2018 Minimizing diseases of peanuts in the Southeastern United States. University of Georgia, University of Florida, Auburn University, Mississippi State University, and Clemson University.
Ketring, D.L 1986 Physiological response of groundnut to temperature and water deficits breeding implications. Agrometeorology of Groundnut. Proceeding of an International Symposium 135-143.
Kvien, C.K., C.C Holbrook, P Ozias-Akins, C Pilon, A.K Culbreath, and T.B Brenneman 2019 Peanut production guide 2019: Peanut Physiology University of Georgia In press.
S.K Leong, and C.K Ong (1983). The influence of temperature and soil water deficit on the development and morphology of peanuts (Arachis hypogaea L.). J. Exp. Bot 34: 1551- 1561.
H.K Lichtenthaler, and A.R Wellburn (1983). Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans 11: 591- 592.
J. D Maguire, (1962). Speed of germination-aid selection and evaluation for seedling emergence and vigor. Crop Sci 2: 176- 177.
K Maxwell, and G.N Johnson (2000). Chlorophyll fluorescence-a practical guide. J. Exp. Bot 51: 659- 668.
C.K Ong, (1984). The influence of temperature and water deficit on partitioning of dry matter in groundnut (Arachis hypogaea L.). J. Exp. Bot 35: 746- 755.
Pilon, C., F Bourland, and D Bush 2016 Seeds and Planting Pages 67- 83 In Linking Physiology to Management The Cotton Foundation, Cordova, TN.
C., Pilon, J.L Snider, V Sobolev, D.R Chastain, R.B Sorensen, C. D Meeks, A.N Massa T Walk, B Singh, and H. J Earl (2018). Assessing stomatal and non-stomatal limitations to carbon assimilation under progressive drought in peanut (Arachis hypogaea L.). J. Plant Physiol 231: 124- 134.
Porter, W.M 2019 Water utilization and irrigation management. In University of Georgia Peanut Production Quick reference guide University of Georgia, GA http://www.gapeanuts.com/growerinfo/2019_ugapeanutguide.pdf. (accessed 06 April 2019) .
P.V.V., Prasad, K.J Boote, J.M.G Thomas, L.H Allenand D.W Gorbet (2006). Influence of soil temperature on seedling emergence and early growth of peanut cultivars in field conditions. J. Agron. Crop Sci 192: 168- 177.
S.K Ries, and E.H Everson (1973). Protein content and seed size relationship with seedling vigor of wheat cultivars. Agron. J 65: 884- 886.
M.E Salvucci, and S.J Crafts-Brander (2004). Inhibition of photosynthesis by heat stress: The activation state of rubisco as a limiting factor in photosynthesis. Physiol 78: 619- 622.
S.M., Schrader R.R Wise, W.F Wacholtz, D.R Ort, and T.D Sharkey (2004). Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant Cell Environ 27: 717- 724.
J.L., Snider, G.D Collins, J Whitaker, K.D Chapman, P Horn, and T.L Grey (2014). Seed size and oil content are the key determinants of seedling vigor in Gossypium hirsutum. J. Cotton Sci 18: 1- 9.
J.L., Snider, G.D Collins, J Whitaker, K.D Chapman, and P Horn (2016). The impact of seed size and chemical composition on seedling vigor, yield and fiber quality of cotton in five production environments. Field Crops Res 193: 186- 195.
Taiz, L and E Zeiger 2010 Plant Physiology. 5th ed Sinauer Associates, Massachusetts.
R.S Tubbs, (2019). Peanut crop rotations. In University of Georgia Peanut Production Quick reference guide. GA: University of Georgia. , .
D.F., Wanjura, E.B Hudspethand J.D Bilbro (1969). Emergence time, seed quality and planting depth effects on yield and survival of cotton (Gossypium hirsutum L.). Agron. J 61: 63- 65.
Williams, J.H and K.J Boote 1995 Physiology and modeling predicting the unpredictable legume Pages 301- 353 In: Advances in Peanut Science American Peanut Research and Education Society, Stillwater, OK.
R.R., Wise, A.J Olson, S.M Schrader, and T.D Sharkey (2004). Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ 27: 717- 724.
I.M.W Wood, (1968). The effect of temperature at early flowering on growth and development of peanuts. Aust. J. Agr. Res 19: 241- 251.
Notes
- First, second, and third authors: Graduate Student, Assistant Professor, and Associate Professor, Department of Crop and Soil Sciences, University of Georgia, 2360 Rainwater Road, Tifton, GA 31793. [^] *Corresponding author's E-mail: cpilon@uga.edu
Author Affiliations