

Introduction
Peanut (Arachis hypogaea L.) is widely cultivated across the United States and throughout the world for its edible seeds which are profitable to producers and nutritious for consumers. The United States produces the third largest peanut crop in the world, behind China and India, reporting a 2015 crop of 6 billion pounds valued at $1.2 B (National Agricultural Statistics Service (NASS), 2016). Of the four peanut market-types grown in the U.S., only two (runner and Virginia) are grown nation-wide. Spanish-types are grown only is Southwestern U.S. regions of Oklahoma and Texas, while Valencia-types are primarily grown in New Mexico and the Texas high-plains region. Valencia peanuts (Arachis hypogaea L. subsp. fastigiata var. fastigiata), known for their 3-4 seeded pods, red seed coat, and sweet flavor, are sold mainly in specialty markets such as organic or all-natural and are usually marketed in-shell or as peanut butter. Although Valencia peanuts account for less than one percent of U.S. production, they make up the majority of the U.S. organic peanut market.
Certified organic peanut production is complex and highly regulated. Peanut is susceptible to a number of stem, foliage, pod, and root diseases (Melouk and Backman, 1995). Since peanut plants form their seed underground, crop productivity is often threatened by soil-borne pathogens such as fungi. Soil-borne fungi cause diseases that adversely affect peanut health, productivity, and seed quality throughout all growing areas of the United States. Pod rot (Rhizoctonia solani Kühn, Pythium myriotylum), crown rot (Aspergillus niger Teigh), and southern blight (Sclerotium rolfsii Sacc) occur in all U.S. peanut-producing areas, while others such as Sclerotinia blight (Sclerotinia sp.) are limited to certain geographic regions.
First observed in the U.S. in Virginia in 1971, Sclerotinia blight is now a major concern to peanut producers in the Southwest US (Oklahoma and Texas), Virginia, and North Carolina. The disease is characterized by wilting and yellowing of branches (flagging) as well as stem bleaching and shredding above ground (Melouk and Backman, 1995). Depending upon severity of field infestation, yield losses due to Sclerotinia blight are typically 10% but may be as high as 50% (Melouk and Backman, 1995). The disease can be caused by two species of Sclerotinia, Sclerotinia minor Jaggar and Sclerotinia sclerotiorum (Lib) de Bary (Porter and Melouk, 1997), with the S. minor being more prevalent in the United States, and S. sclerotiorum more common in Australia (Cruickshank, et al., 2002) and Argentina (Marinelli, et al., 1998). Peanuts with resistance to S. minor have also been shown to be resistant to S. sclerotiorum (Cruickshank et al., 2002, Woodward et al., 2006). Sclerotinia blight (S. sclerotiorum) has been reported in Georgia (Woodward et al., 2006), Nebraska (Melouk et al., 2003), and on Valencia peanut in eastern New Mexico (Sanogo and Puppala, 2007; Lujan et al., 2016). The extent to which S. sclerotiorum has invaded New Mexico is unknown, but Sclerotinia blight is wide-spread in Texas and the percentage of the disease caused by S. sclerotiorum versus S. minor is unknown. The sclerotial form of both pathogens may persist in soil for many years and can be spread by unclean production equipment or avian waste. Expensive fungicides applied throughout the growing season are required for effective Sclerotinia blight control and are not an option for the organic Valencia peanut producers in New Mexico. Host plant resistance will provide the most effective solution to managing Sclerotinia blight in any peanut production system, but is essential to organic production. Several Valencia cultivars have been released for production including New Mexico Valencia A (Hsi and Finkner, 1972), New Mexico Valencia C (Hsi, 1980), GenTex 101, 102, and 136 (Borden Peanut Company, Portales, NM), Georgia Red (Branch and Hammons, 1987), Georgia Valencia (Branch, 2001), and NuMex01 (Puppala and Tallury, 2014), but none have documented levels of resistance to Sclerotinia blight.
Limitations to successful breeding of Sclerotinia blight resistant peanut cultivars are numerous and include the quantitative nature of the trait (Wildman et al., 1982) along with the narrow genetic background of cultivated peanut (Simpson et al., 2001) and the lack of identified resistance sources. Little progress has been made in identifying sources of Sclerotinia blight resistance in Valencia-type peanut (Smythe et al., 2011), despite the development of a Valencia germplasm core collection (Dwivedi et al., 2008). Phenotyping peanut germplasm collections for specific traits in extremely useful in revealing sources of disease resistance. Examination of the U.S. peanut germplasm collection has led to the identification of resistance to leafspot (Holbrook and Anderson, 1995), Rhizoctonia limb rot (Franke et al., 1999), tomato spotted wilt virus (Wang et al., 2007), root-knot nematode (Holbrook et al., 2000), Sclerotinia blight (Damicone et al., 2010), and pepper spot (Damicone et al., 2010). With the development of molecular markers associated with phenotypic traits, it is possible to screen large germplasm collections without years of field evaluation. Genotyping peanut germplasm collections has resulted in the identification of accessions with possible resistance to Sclerotinia blight (Chamberlin et al., 2010; Chamberlin, 2014; Engin et al., 2015). Coupling genotype and phenotype data can result in new sources of disease resistance being incorporated into cultivar releases (Chamberlin et al., 2018). One disadvantage to screening composite germplasm collections is that all peanut market-types are included. For example, only 22% and 15% Valencia accessions are found in the U.S. and ICRISAT core collections, respectively. This study examines the genotype of the Valencia core collection composed of 77 accessions (Dwivedi et al., 2008) using a SSR marker associated with Sclerotinia blight resistance (Chenault et al., 2009).
Materials and Methods
Plant Materials
A total of 77 accessions from the Valencia peanut mini-core collection (Table 1; Dwivedi et al., 2008) were examined in this study. Sclerotinia blight resistant accession PI 274193 and susceptible cultivar Okrun were also included for reference.
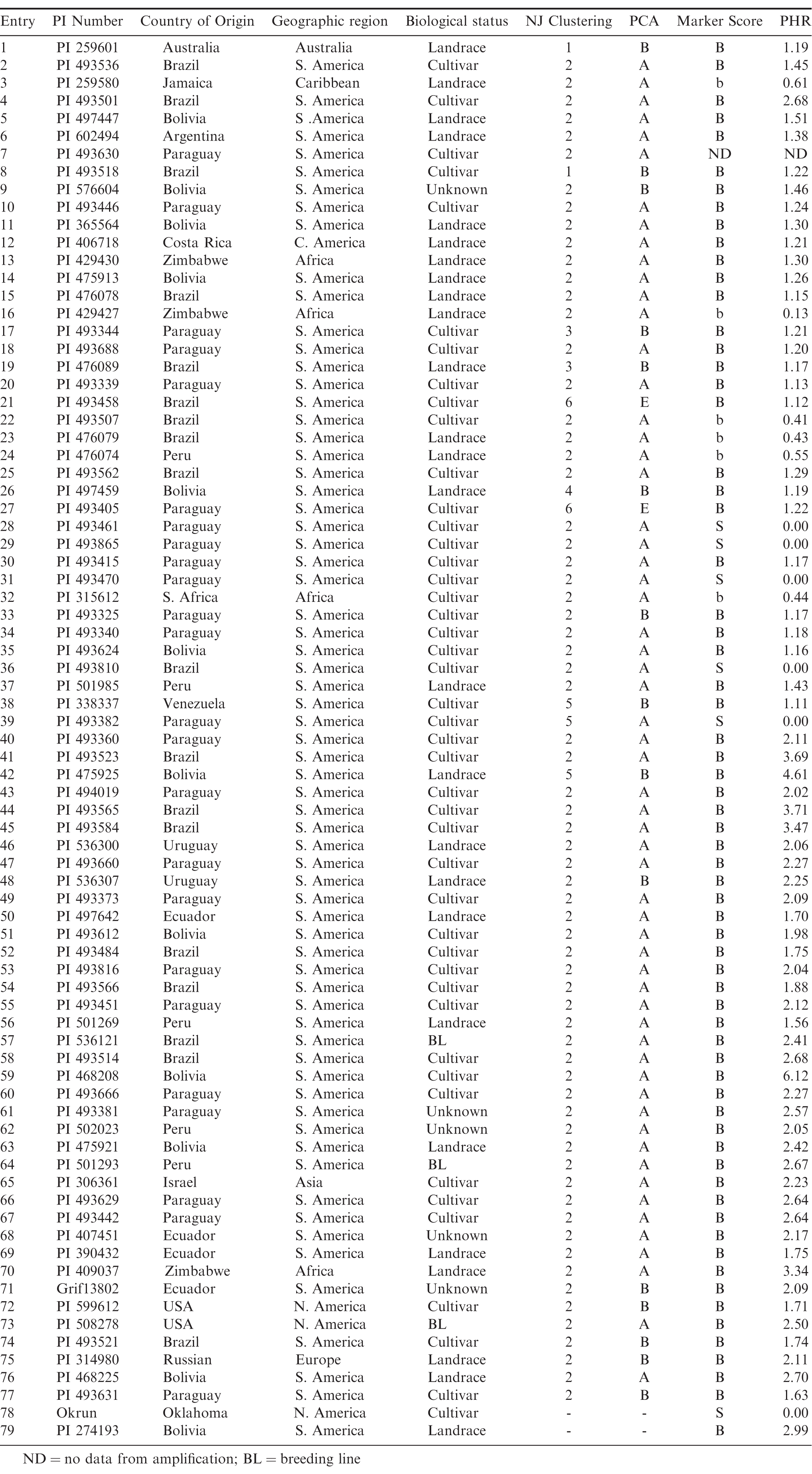
Information for the 77 valencia peanut mini-core accessions genotyped with the SSR marker associated with Sclerotinia blight resistance along with respective Neighbor-Joining Clustering and Principal Coordinate Analysis (PCA) (from Kottapalli et al., 2011), marker score (Chenault et al., 2009) and peak-height ratio (PHR) values.
DNA Extraction and Fragment Analysis
For each plant accession, DNA was extracted from young leaf tissue using the Qiagen Plant DNeasy DNA extraction kit (Qiagen, Germantown, MD), and quantified using a NanoDrop nd-1000 spectrometer (ThermoFisher Scientific) or the Pico Green dsDNA Assay kit (ThermoFisher Scientific). DNA concentrations were adjusted to 25 ng/ul prior to PCR amplification. Amplification was performed in triplicate using an SSR marker derived from SSR primer pair pPGPseq2E6, which has been reported to be associated with Sclerotinia blight resistance in peanut (Chenault et al., 2009). Reaction components: 10 μl (2.5 ng/μl) genomic DNA, 2 μl 10 μl 10X PCR Buffer, 2 μl 25 mM MgCl2,1 μl each 10 mM primers, 2 μl 2 mM dNTP mix, 0.5 μl HotStar Taq DNA Polymerase (5 U/μl; Qiagen), 1.5 μl sterile H2O. Primer sequences are as follows: pPGPseq2E6L (5' TACAGCATTGCCTTCTGGTG 3') and Marker 3 (5' GCACACCATGGCTCAGTTATT 3'). Primers were labeled with 5-FAM fluorophor. Amplification was carried out in a MJ Research PTC-100 thermal cycler under the following conditions: 95 C 15 min.; 34 cycles of 94 C 45 sec., 50 C 1 min., 72 C 90 sec.; 72 C 10 minutes. Fragment analysis of PCR products was done using an Applied Biosystems 3730 DNA Analyzer and sized using a LIZ labeled size standard (ThermoFisher Scientific). Amplification with this primer set generally produces two bands of interest, one at 100 bp (predominant in susceptible genotypes) and one at 115 bp (predominant in resistant genotypes). Sequencing of the amplified bands has shown the molecular basis for the size difference to be the length of the CT repeat sequence. Marker profiles were scored using previously reported methods (Chenault et al., 2009). The ratio of the peak height (PHR) of these two bands serves as a predictor of potential resistance to Sclerotinia blight. Peak height of bands were analyzed using PeakScanner 1.0 software (ThermoFisher Scientific).DNA from susceptible cultivar Okrun and resistant accession PI 274193 were included in each assay. Correlation analysis of Neighbor-Joining (NJ)-clustering and Principal Coordinate Analysis (PCA) taken from Kottapalli et al., 2011, and of genotypic marker data was conducted using PROC CORR, SAS ver. 9.3, Cary, NC.
Results and Discussion
Phenotypic evaluation of peanut germplasm collections previously genotyped with the SSR marker used in this study has confirmed that the presence of the 115 bp band generated during amplification is associated with increased resistance to Sclerotinia blight, and the intensity of that band is directly correlated with the level of resistance seen in the field (Chenault et al., 2009; Chamberlin et al., 2010; Chamberlin and Bennett, in press). Amplification profiles for 86% of the accessions tested in this study consisted of both the 115 bp and 110 bp bands and received a visual marker score of 'B' (115 bp band more intense) when separated on agarose gels. The remaining accession profiles were scored as either 'b' (110 bp band more intense) or 'S' (110 bp band only). Fragment analysis using the same primers labeled with 5-Fam fluorophor allowed more accurate quantification of products than agarose gel visualization, and intensity of each band was estimated by measurement of peak heights. The peak height (PH) ratio of the two bands (PH 115 bp/PH 110 bp) has been shown to be negatively correlated with disease resistance and serves as a predictor of expected resistance levels (r = - 0.68, Chamberlin and Bennett, in press). Accessions with profiles of PHRs less than 0.757 demonstrate disease incidence levels above 15% (High or H), where as those with a PHR between 0.757 and 2.0 generally have disease incidence levels between 5-15% (Moderate or M). Accessions with PHRs above 2.0 are the most resistant, with disease incidence levels below 5% (Low or L). The PHRs for all 77 Valencia mini-core accessions are shown in Figure 1 and in Table 1, along with those for susceptible cultivar Okrun and resistant accession PI 274193. The majority of accessions tested (46%) had PHRs placed in the M group, ranging from 1.11 to 1.98. Eleven out of the 77 accessions had PHRs below 0.757, placing them in the H group. In five accessions (entries 28, 29, 30, 36 and 39), amplification produced only the 100 bp band resulting in a PHR of 0.0, the same as susceptible cultivar Okrun which routinely becomes infected with Sclerotinia blight at rates of 48 - 88% under heavy disease pressure (Melouk et al., 2008). The remainder of the accessions had PHRs above 2.0 (39.5%), ranging from 2.04 - 6.12, and were classified as L. Accession PI 468208 (entry 59), a Bolivian cultivar, and accession PI 475925 (entry 42), a Bolivian landrace, exhibited the highest PHRs at 6.12 and 4.61, respectively. In total, 30 accessions with PHRs above 2.0 were placed in the "L group" and would be expected to demonstrate less than 5% disease incidence in the field.
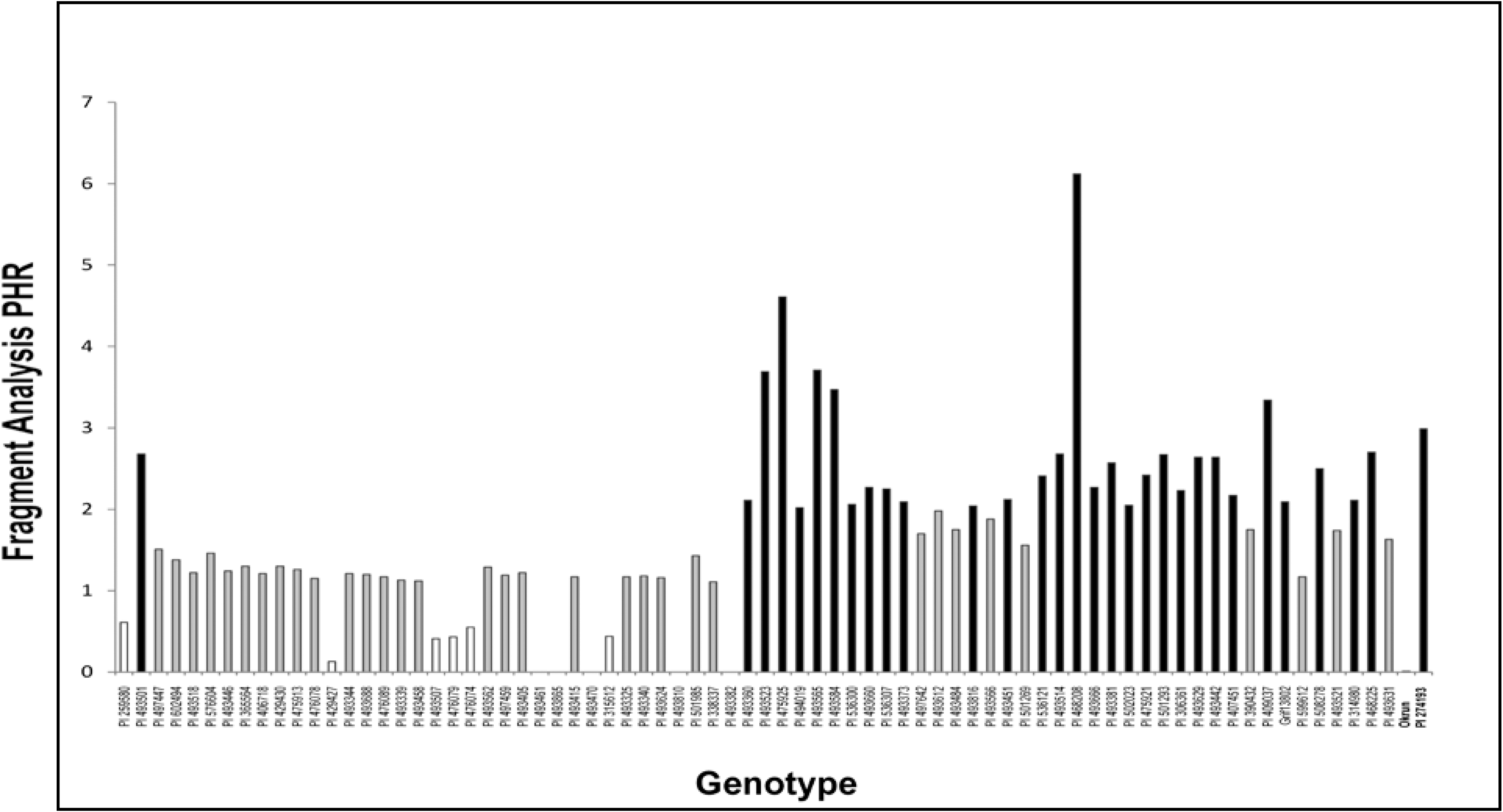
Peak-height ratios (PHRs) for all 77 Valencia mini-core accessions tested, along with those for susceptible cultivar Okrun and resistant accession PI 274193.
Kottapalli et al. (2011) examined the population structure and diversity of the Valencia core collection with 52 SSR loci and separated the accessions into 6 clusters and two distinct major groups using Neighbor-joining (NJ) clustering, principal coordinate analysis (PCA) and STRUCTURE analysis (Table 1). Pearson correlation analysis (PROC CORR, SAS, ver. 9.3) of that data along with the marker data collected here for Sclerotinia blight resistance showed no correlation of potential resistance and accession cluster. Furthermore, no correlation of potential resistance and geographical origin was observed.
Sclerotinia diseases cause significant losses annually in a broad range of crop species. Some of the greatest losses occur in cropping environments that are optimal for plant growth, such as irrigated fields, where a lush, dense plant canopy is present. Sclerotinia sclerotiorum has been reported to pose a significant threat to peanut production in Argentina (Marinelli et al., 1998) and Australia (Cruickshank et al., 2002) and has also been reported in Rhodesia (Rothwell, 1972) and China (Yan et al., 2014). The pathogen was reported on peanut in the U.S. in the Virginia-Carolina region (Beute et al., 1975), and appears to be spreading across peanut producing states as it has more recently been reported in Georgia (Woodward et al., 2006),Texas, (Woodward et al., 2008), and New Mexico (Sanogo and Puppala, 2007; Lujan et al., 2016). Although Sclerotinia minor Jagger is most commonly associated with Sclerotinia blight on peanut in the Southwestern U.S., mixed infections of S. minor and S. sclerotiorum are possible and have been reported on vegetable crops (Kim and Cho, 2002), and on sunflower (Sedun and Brown, 1989;Tozlu and Demirci, 2008).
The recent report of Sclerotinia blight caused by S. sclerotiorum on peanut in New Mexico and Texas has caused concern that the disease may eventually pose a threat to organic Valencia peanut production in that region. Since certified organic production prohibits the use of chemical pesticides, cultivars with physiological resistance to Sclerotinia spp. offer the most inexpensive and sustainable solution for disease management. Few have reported on the study of peanut resistance to S. sclerotiorum (Porter et al., 1975; Coffelt 1980; Cruickshank, et al., 2002), but several cultivars have been released in the last decade with varying levels of resistance to S. minor (Isleib et al., 2011; Baring et al., 2013; Melouk et al., 2013; Simpson et al., 2013; Chamberlin et al., 2015; Isleib et al., 2015; Chamberlin et al., in press).
The use of marker assisted selection in cultivar breeding not only expedites screening germplasm collections for disease resistance but also leads to more efficient advanced breeding line development. To date no molecular marker associated with S. sclerotiorum resistance in peanut has been reported, but Vuong et al. (2008) reported genotyping a population of recombinant inbred lines (RILs) derived from a cross with a partially resistant plant introduction (PI) which led to the identification of QTL associated with resistance in soybean. There is evidence that the mechanisms of resistance to the two pathogens are similar in peanut and other crop species, and that genotypes resistant to S. minor may also express that resistance against S. sclerotiorum (Sedun and Brown, 1989; Cruickshank et al., 2002). Therefore, it is possible that screening germplasm collections using a marker associated with S. minor resistance would be effective in the identification of accessions with potential resistance to S. sclerotiorum as well, and this possibility is being explored. In this study, such screening led to the elimination of 61% of the Valencia core collection as candidates for excellent resistance to Sclerotinia blight, thus reducing time and space required for field phenotyping this collection for that specific trait. Although validation by field testing will be necessary, the results of this study predict 30 accessions from the Valencia core collection will be highly resistant to Sclerotinia blight and should be considered as potential sources of resistance for breeding programs focused on the Valencia market-type.
Acknowledgements
The investigators would like to thank Lisa Myers for technical assistance. This research was supported by USDA-ARS CRIS Project No. 3072-21220-007-00D. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Literature Cited
M.R., Baring, C.E. Simpson, M.D. Burow, J.M. Cason, and J.L. Ayers (2013). Registration of 'TamrunOL11' peanut. J. Plant Reg 7: 154- 158.
W.D., Branch, and R.O. Hammons (1987). Registration of Georgia Red peanut. Crop Sci 27: 1090.
W.D. Branch, (2001). Registration of Georgia Valencia. Crop Sci 41: 2002- 2003.
M.K., Beute, D.M. Porter, and B.A. Hadley (1975). Sclerotinia blight of peanut in North Carolina Virginia and its chemical control. Plant Dis. Rep 59: 697- 701.
K.D. Chamberlin, (2014). Characterization of ICRISAT peanut mini core accessions with regards to a molecular marker associated with resistance to Sclerotinia blight. Peanut Sci 41: 42- 49.
Chamberlin, K.D., and R.S. Bennett (in press). Screening of the U.S. peanut mini-core collection for resistance to Sclerotinia blight Proc. Am. Peanut Res. Ed. Soc 49 (abstr.).
K.D., Chamberlin, R.S. Bennett, and J.P. Damicone ((2018)). Registration of 'Lariat' peanut. J. Plant Reg 12: 36- 42.
K.D., Chamberlin, R.S. Bennett, J.P. Damicone, C.B. Godsey, H.A. Melouk, and K. Keim (2015). Registration of 'OLé' peanut. J. Plant Reg 9: 154- 158.
K.D.C., Chamberlin, H.A. Melouk, and M.E. Payton (2010). Evaluation of the U.S. peanut mini core collection using a molecular marker for resistance to Sclerotinia minor Jagger. Euphytica 172: 109- 115.
K.D., Chenault, A. Mass, J.P. Damicone, M.E. Peyton, and H.A Melouk (2009). Discovery and characterization of a molecular marker associated with Sclerotinia blight. Euphytica 166: 357- 365.
T.A. Coffelt, (1980). Registration of VGP 1 peanut germplasm. Crop Sci 20: 419.
A.W., Cruickshank, M. Cooper, and M.J. Ryley (2002). Peanut resistance to Sclerotinia minor and S. Sclerotiorum. Aust. J. Agric. Res 53: 1105- 1110.
J.P., Damicone, C.C. Holbrook, D.L. Smith, H.A. Melouk, and K.D. Chamberlin (2010). Reaction of the core collection of peanut germplasm to Sclerotinia blight and pepper spot. Peanut Sci 37: 1- 11.
S.L., Dwivedi, N. Puppala, H.D. Upadhyaya, M. Manivannan, and S. Singh (2008). Developing a core collection of peanut specific to the Valencia market type. Crop Sci 48: 625- 632.
Y., Engin, H.D. Upadhyaya, and U. Bulent (2015). Molecular diagnosis to identify new sources of resistance to Sclerotinia blight in groundnut (Arachis hypogaea L.). Euphytica 203: 367- 374.
M.D., Franke, T.B. Brenneman, and C.C. Holbrook (1999). Identification of resistance to Rhizoctonia limb rot in a core collection of peanut germplasm. Plant Dis 83: 944- 948.
D.C. Hsi, (1980). Regisration of New Mexico Valencia C. Crop Sci 20: 113- 114.
D.C., Hsi, and R.E. Finkner (1972). Registration of New Mexico Valencia A. Crop Sci 12: 256.
C.C., Holbrook, and W.F. Anderson (1995). ., Evaluation of a core collection to identify resistance to late leafspot in peanut. Crop Sci 35: 1700- 1702.
C.C., Holbrook, P. Timper, and H.Q. Xue (2000). Evaluation of the core collection approach for identifying resistance to Meloidogyne arenaria in peanut. Crop Sci 40: 1172- 1175.
T.G., Isleib, S.R. Milla-Lewis, H.E. Pattee, S.C. Copeland, M.C. Zuleta, B.B. Shew, J.E. Hollowell, T.H. Sanders, L.O. Dean, K.W. Hendrix, M. Balota, and J.W. Chapin (2011). Registration of 'Bailey' peanut. J. Plant Reg 5: 27- 39.
T.G., Isleib, S.R. Milla-Lewis, H.E. Pattee, S.C. Copeland, M.C. Zuleta, B.B. Shew, J.E. Hollowell, T.H. Sanders, L.O. Dean, K.W. Hendrix, M. Balota, J.W. Chapin, and M.S. Monfort (2015). Registration of 'Sugg' peanut. J. Plant Reg 9: 44- 52.
W.G., Kim, and W.D. Cho (2002). Occurrence of Sclerotinia rot on composite vegetable crops and the causal Sclerotinia spp. Mycobiology 30: 41- 46.
P., Kottapalli, H.D. Upadhyaya, K.R. Kottapalli, P. Payton, S. Dwivedi, M. Burow, K.O. David, S. Sanogo, and N. Puppala (2011). Population structure and diversity in Valencia peanut germplasm collection. Crop Sci 51: 1089- 1100.
P., Lujan, S. Sanogo, N. Puppala, and J. Randall (2016). Factors affecting mycelium pigmentation and pathogenicity of Sclerotinia sclerotiorum on Valencia peanut. Can. J. Plant Sci 96: 461- 473.
A., Marinelli, G.J. March, A. Rago, and J. Giuggia (1998). Assessment of crop loss in peanut caused by Sclerotinia sclerotiorum, S.minor and Sclerotium rolfsii in Argentina. Inter. J. Pest. Man 44: 251- 254.
H.A., Melouk, K.D. Chamberlin, C.B. Godsey, J. Damicone, M.D. Burow, M.R. Baring, C.E. Simpson, K.E. Dashiell, and M.E. Payton (2013). Registration of 'Red River Runner' peanut. J. Plant Reg 7: 22- 25.
Melouk, H.A., K.D. Chenault, J.P. Damicone, and C.B. Godsey 2008 Sclerotinia blight resistance in peanuts Oklahoma Ag. Exp. Sta. Rep. Partners in Progress: Peanuts P-1019.
Melouk, H.A., K.E. Jackson, and J.P. Damicone 2003 First report of Sclerotinia blight on peanut in Nebraska Phytopathology 93:S76 (abstr.).
Melouk, H.A., and P.A. Backman 1995 pp. 75-85 In: Melouk HAand Shokes FM (eds.) Peanut Health Management, APS Press, St. Paul, Minnesota.
National Agricultural Statistics Service (NASS), Agricultural Statistics Board, United States Department of Agriculture (USDA), (2016) Crop Production Report, ISSN: 1936-3737.
D.M., Porter, M.K. Beute, and J.C. Wynne (1975). Resistance of peanut germplasm to Sclerotinia sclerotiorum. Peanut Sci 2: 78- 80.
Porter, D.M., and H.A. Melouk 1997 Sclerotinia blight In: Compendium of Peanut Diseases N. Kokalis-Burelle, D.M. Porter, R. Rodriquez-Kabana, D.H. Smith, and P. Subrahmanyam, eds., American Phytopathological Society, St. Paul, MN pp 34- 6.
N., Puppala, and S.P. Tallury (2014). Registration of NuMex 01 high-oleic Valencia peanut. J. Plant Reg 8: 127- 130.
A. Rothwell, (1972). Groundnut diseases in Rhodesia. Rhodesia Ag. J 69: 35- 38.
S., Sanogo, and N. Puppala, (2007). Characterization of a darkly-pigmented mycelial isolate of Sclerotinia sclerotiorum on Valencia peanut in New Mexico. Plant Dis 91: 1077- 1082.
F.S., Sedun, and J.F. Brown (1989). Comparison of three methods to assess resistance in sunflower to basal stem rot caused by Sclerotinia sclerotiorum and S. minor. Plant Dis 73: 52- 55.
C.E., Simpson, A. Krapovickas, and J.M. Valls (2001). History of Arachis including evidence of A. hypogaea progenitors. Peanut Sci 28: 79- 81.
C.E., Simpson, J.L. Starr, M.R. Baring, M.D. Burow, J.M. Cason, and J.N. Wilson (2013). Registration of 'Webb' peanut. J. Plant Reg 7: 265- 268.
Smythe, B., S. Sanogo, N. Puppala, S. Thomas, and R. Steiner 2011 Screening of a Valencia peanut core collection for resistance to Sclerotinia sclerotiorum Phytopathology 101:S168; (abstr.).
E., Tozlu, and E. Demirci (2008). Incidence and characterization of sunflower stem rot disease caused by Sclerotinia sclerotiorum and S. minor in Pasinler Plan of Erzurum, and reaction of some sunflower cultivars to the pathogens. Bitki koruma Bul 48: 19- 33.
T.D., Vuong, B.W. Diers, and G.L. Hartman (2008). Identification of QTL for resistance to Sclerotinia stem rot in soybean plant introduction 194639. Crop Sci 48: 2209- 2214.
M.L., Wang, D.L. Pinnow, and R.N. Pittman (2007). Preliminary screening of peanut germplasm in the US collection for TSWV resistance and high flavonoid content. Plant Path. J 6: 219- 226.
L.G., Wildman, O.D. Smith, C.E. Simpson, and R.A. Taber (1992). Inheritance of resistance to Sclerotinia minor in selected Spanish peanut crosses. Peanut Sci 19: 31- 34.
J.E., Woodward, T.B. Brenneman, R.C. Kemerait, A.L. Culbreath, and J.R. Clark (2006). Sclerotinia blight in Georgia and evidence for resistance to Sclerotinia sclerotiorum in runner peanuts. Plant Health Prog. (May), 1- 7.
J.E., Woodward, C. Nui, R.J. Wright, and M.A. Batla (2008). First report of Sclerotinia sclerotiorum infecting peanut in Texas. Plant Dis 92: 1468.
L.Y., Yan, Y.P. Kang, Y. Lei, J.Q. Huang, L.Y. Wan, and B.S. Liao (2014). First report of Sclerotinia sclerotiorum causing Sclerotinia blight on peanut (Arachis hypogaea) in Northeastern China. Plant Dis 98: 156.
Notes
- USDA-ARS, Wheat, Peanut and other Field Crops Research Unit, 1301 N. Western, Stillwater, OK 74075; [^]
- New Mexico State University, Department of Plant and Environmental Sciences, Clovis, NM, 88102. [^] *Corresponding author: kelly.chamberlin@ars.usda.gov; Ph: 405-624-4141; Fax: 405-624-4142
Author Affiliations