

Introduction
High oleic peanuts have conservatively twice the post-roast shelf life of conventional peanuts as measured by descriptive flavor panels (Braddock et al.1995; Mugendi et al.1998). This improvement results from their unique oil composition, specifically their increased levels of oleic acid, hence the name high oleic, and corresponding decreased levels of linoleic acid (Andersen et al. 1998). This fatty acid profile, richer in monounsaturated fats, is inherently more resistant to oxidation compared to conventional peanuts. In current markets, high oleic peanuts are especially valued in certain confectionary applications in which price points preclude effective packaging oxidation barriers and the products are consumed on a single kernel or multi-kernel basis. However, as just a few rancid (oxidized) kernels can negatively impact the eating experience of the entire product for the consumer, the industry has developed purity requirements around the marketing of high oleic lots. Commonly, 95/100 kernels must pass the minimal fatty acid chemistry threshold (Sweigart et al, 2011), i.e. about 74 oleic acid or greater, and about 8 or lower linoleic acid, with a corresponding O/L ratio of 9.0 or greater (Knauft et al, 2000). Kernels not meeting threshold could be true contamination with conventional peanuts, or the peanuts not meeting threshold could be immature and not fully expressing this high oleic chemistry.
When sampling from kernel to kernel to kernel within a sample of high oleic (or conventional) peanuts, regardless of potential contamination with conventional oleic kernels, a natural distribution in oleic acid/linoleic acid content is observed among the individual kernels. As peanuts mature, it's established that the oleic acid () increases while linoleic acid () decreases, hence the overall O/L ratio increases with maturity (Pattee et al, 1974; Klevorn et al, 2016). However, such changes are poorly understood at the single kernel resolution across populations of interest.
With these two sources of variation in mind, i.e. true contamination of high oleic lots with normal oleic seed, and the natural variation associated with maturity, it's not clear what factors are the predominate cause of commercial lots of high oleic peanuts being sampled and subsequently not meeting established purity limits. It's also important to recognize that measuring this chemistry is both expensive and time consuming. The primary chemical method is gas chromatography (GC) based which requires a skilled operator and 100 extractions/injections of a typical sample. Refractive index methods are developed which correlate well with the primary GC method, and while faster and cheaper than the GC method (Davis et al, 2013), are still relatively time consuming. Near infrared (NIR) methods for measuring this fatty acid chemistry are being implemented in many breeding programs (Chamberlin et al, 2014), but good calibrations on the single seed level for this fatty acid chemistry, especially at high speeds, are challenging.
Terminating flowers generated after about 90 days after planting, which have no potential to generate viable peanuts, has been shown to improve farmer stock yield and grade (Lamb et al, 2017). Late season flower termination also improves maturity distributions, and therefore may improve single kernel oleic acid () distributions.
The objectives of these experiments were to better understand factors influencing single kernel oleic acid () distributions, including seed maturity, seed size, growing environment and/or late season flower termination.
Materials and Methods
Materials
Two high oleic peanut cultivars were planted for this study. The first, 13AU-12 is an advanced runner breeding line from the joint Auburn University/National Peanut Research Laboratory breeding program. The second cultivar, AT-9899 is a high oleic spanish cultivar. Some seed lots of AT-9899 were found to be contaminated with conventional oleic peanuts in 2009. In 2010 work was carried out to recover and purify this line (Chen et al. 2014), and seeds tested in this experiment were from the recovered AT-9899.
NIR Single Seed Analyses
High speed NIR data was collected on individual seed using a Luminar AOTF NIR Spectrometer (SeedMeister), and the Snap32! software package by Brimrose Corporation of America. The raw spectral data was imported into The Unscrambler version 10.3, by CAMO Software, for post-processing and chemometric analysis. A percent-oleic acid prediction result was generated for each seed using a Partial Least Squares (PLS) regression model previously developed and validated for use with spanish and runner peanut seed.
The calibration data set consisted of 378 runner and spanish redskin peanuts ranging from 41 to 83 oleic acid (ratio in oil fraction of the peanut). The PLS model was developed by pairing the spectral data from each kernel with its corresponding oleic acid result from Fatty Acid Methyl Ester (FAME) analysis by gas chromatography. Analysis of an independent validation set of seeds resulted in a Root Mean Squared Error (RMSE) of 4.4. The uncertainty of the prediction results could then be estimated to be 9 oleic acid at a 95 confidence interval (CI). Later in the process of data collection, a second validation was performed to include selected seeds from the study to verify that the calibration accuracy was being maintained. The RMSE was 4.3 for the verification, which was the same as the initial validation set.
Prior to planting, seeds were screened via NIR. Only seed measuring greater than 75 oleic acid were saved for planting. A total of 2189 spanish and 2648 runner seeds were screened as summarized in Table 1. For this, 98.9 and 94.1 of spanish and runner seed, respectively, met specified criteria for planting. Given the RMSE of the calibration model it was assumed all peanuts planted met the common industry accepted threshold of 74 oleic acid or an O/L ratio 9.0.
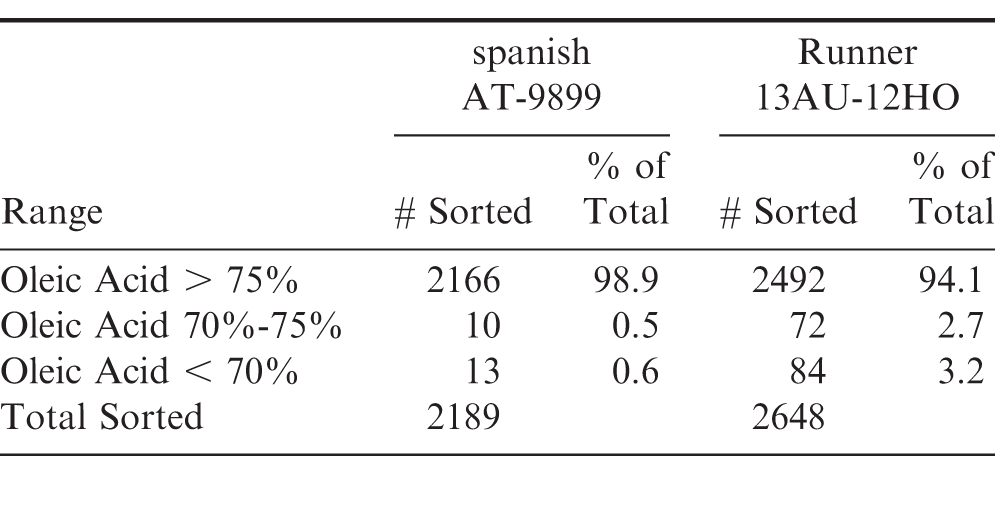
Summary of NIR sorting of individual runner and spanish seed prior to planting. Only seed sorted with an oleic acid content greater than 75 as measured by NIR were planted for the current study.
After planting, harvesting, sorting, and shelling as described below, individual redskin peanuts were scanned via NIR to measure the percent oleic acid. Sample data was collected for each maturity or size subset by scanning 100 seed from labelled bags. For some kernel classifications such as the yellows or No1-sized, 100 kernels were not available but all kernels available were measured. For the harvested samples sorted by maturity and size, a total of 12,426 and 11,697 kernels, respectively, were scanned via NIR. Kernels were randomly hand-selected, after discarding only the ones that were not suitable for NIR analysis due to extremely small size or defects.
Each unique 100-kernel sample set was given a descriptive filename corresponding to its organizational grouping (eg Yellow No 1). After completing data collection and processing, the numbered and categorized prediction results were organized into an Excel spreadsheet for further analysis. Excel and JMP Pro 12.0.1 of SAS Institute Inc. were used for statistical data analysis. Treatment means were compared using the Tukey-Kramer HSD Test.
Planting, harvesting, sorting and shelling
Runner and spanish peanuts screened having more than 74 oleic acid were planted at a seeding rate of 20 seed/m in environmental control plots located at the USDA ARS NPRL in Dawson GA on 26 May 2014. Soil type was a Tifton sandy loam (Fine-loamy, kaolinitic, thermic Plinthic Kandiudults). Experimental units were 0.91-m rows within the environmental control plots. The average plant population after emergence was approximately 17.1 plants/m for both seed types. Plants were fully irrigated based on soil moisture sensing (MPS-2, Decagon Devices, Pullman, WA). Soil nutrition, fungicides, herbicides, and insecticides were applied throughout the growing season in accordance with recommended production practices. Soil temperature adjustments and hand flower removal were initiated approximately 90 days after planting (DAP) on 18 Aug 2014 as described in the following paragraph. Spanish peanuts were dug 122 days after planting on 15 Sep 2014. Runner peanuts were dug 138 days after planting on 30 Sep 2014.
Treatments for both spanish and runner peanuts included three soil temperature treatments: ambient (untreated control), heated or chilled; two late season flower terminations: control or hand removal. All experimental units were replicated 4 times. The schematic of the plot layout is shown in Figure 1.
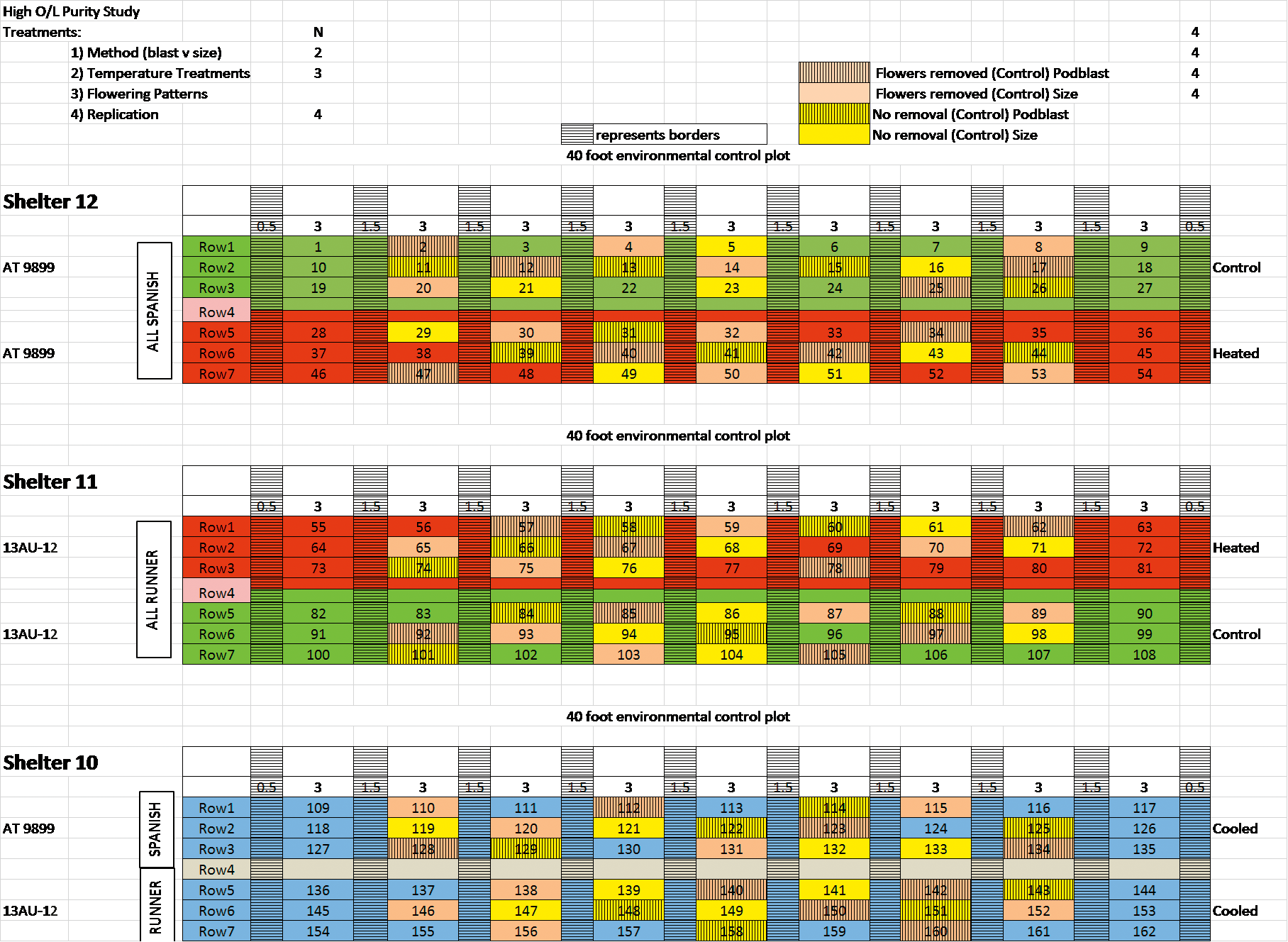
Schematic of environmental test plots and experimental design used in the current study.
Heating cables and chilled water coils were used to heat and cool, respectively, the soil in the pod zone portions of the environmental control plots (Figure 1). Type T thermocouples were installed in each soil zone (untreated, heated, and cooled). A microprocessor (CR1000, Campbell Scientific, Inc., Logan, UT) monitored the average soil temperature in each zone. The heating cables were controlled by the microprocessor so that the heated soil temperature was maintained 4.4C above the average soil temperature of the untreated control. The microprocessor was programmed to control the flow of chilled water through the cooling coils to maintain the soil 4.4C below that of the untreated control, but the chiller ran continuously and was only able to maintain the soil 2.8C below the ambient soil temperature. The average soil temperature for each of the soil temperature treatments over a typical week of implementation are shown in Figure 2.
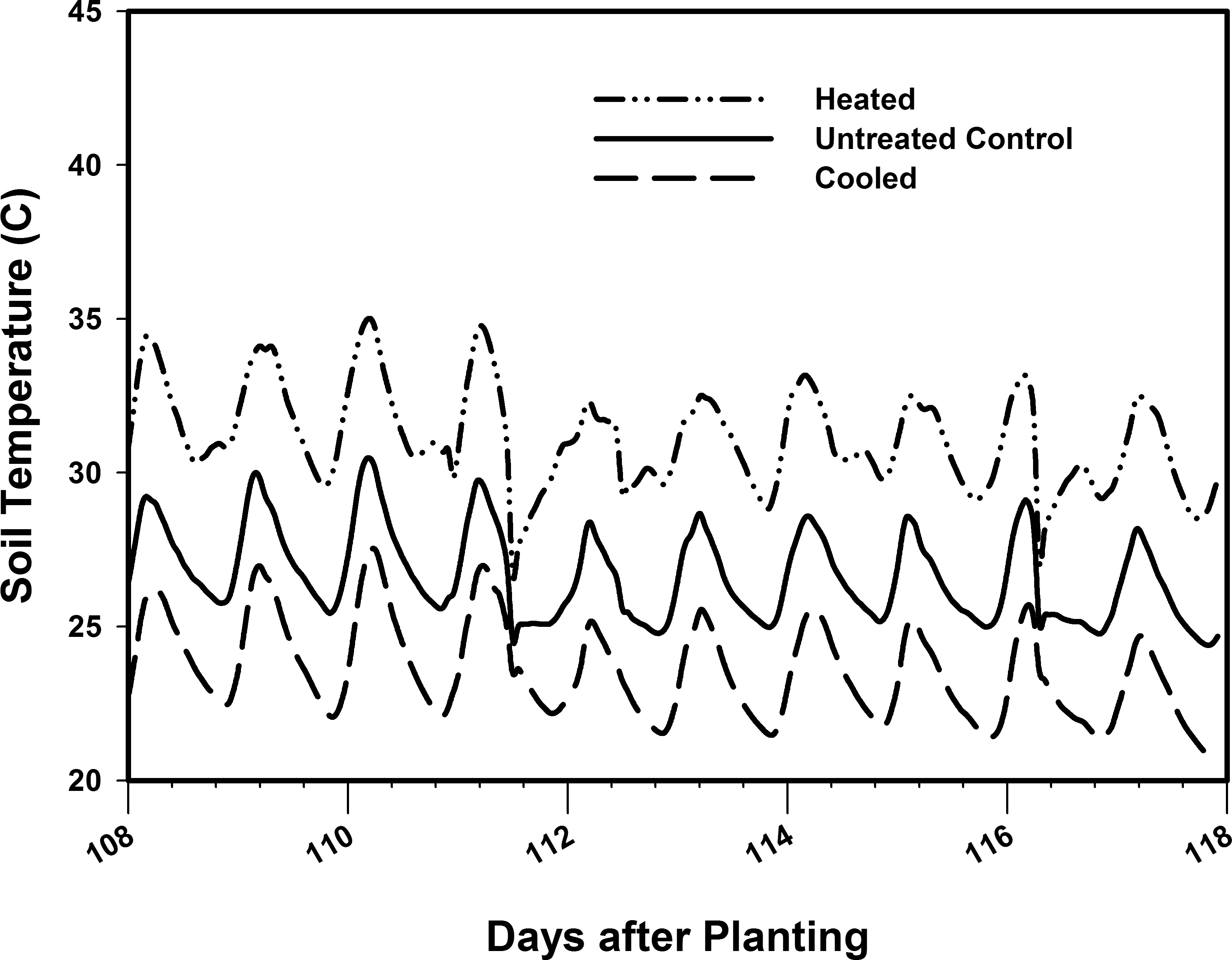
Pod zone temperatures (C) measured for 1 week in environmental control plots.
Late season flowers, i.e. flowers added by the peanut plants after about 90 days, were hand removed for select treatments. Trained personnel hand removed flowers each morning between 6:30 and 8:00 AM by visually looking for and pinching off new flowers.
At harvest, plots were split and half the material was processed to simulate common, best commercial processing while the other half was processed for maturity sorting. Peanuts experimentally processed to simulate industrial practices, by hand digging, then inverting in windrows, and field curing for approximately 3 days. After windrow curing, peanuts were harvested with a plot thresher (Kingaroy Engineering Works, Kingaroy, Queensland, AU). Peanuts were mechanically cured until the kernel moisture content was less than or equal to 10 using air heated 8C above ambient, but no greater than 35C in laboratory-scale dryers (Butts, et al., 2002). Peanuts were shelled using a Model 4 sample sheller (Davidson and McIntosh, 1973) and kernels sorted into APSA standard size grades, including Jumbo, Medium and No1 for runners; Jumbo and No1 for spanish peanuts. For runner peanuts, Jumbo-sized peanuts did not pass an 8.3 mm by 19.1 mm slotted screen, the Medium-sized peanuts fell through the Jumbo screen but did not pass a 7.1 mm by 19.1 mm slotted screen, and No1-sized peanuts fell through the Medium screen but did not pass a 6.4 mm by 19.1 mm slotted screen. For spanish peanuts, Jumbo-sized peanuts did not pass a 6.4 mm round screen, while the No1-sized peanuts fell thru the Jumbo screen but did not pass a 5.9 mm by 19.1 mm slotted screen.
The remaining peanuts were hand dug, turned, loose soil shaken off, and pod blasted to remove the exocarp (Williams and Drexler, 1981). Trained personnel subsequently sorted and bagged peanuts according to exposed mesocarp color for drying and shelling. Maturity categories included black, brown, orange and yellow.
Results and Discussion
Half the test plots from this study were harvested, handled and shelled to simulate common, best industrial practices, including windrow drying, mechanical curing, and shelling/sizing according to industry standards. The other half were immediately pod blasted on the day of harvest, sorted according to maturity based on the exposed mesocarp color, mechanically dried, and then shelled. These two portions of the study are referred to as simulated commercial processing and maturity sorting. The current manuscript primarily focuses on plots processed via simulated commercial processes, although data generated via maturity sorting is used to complement findings.
Total shelled material recovered per 0.9 m of experimental row, defined as the total mass of Jumbo's, No1's, sounds splits and oilstock for spanish, or the total mass of Jumbo's, Medium's, No1's, sound splits and oilstock for runners, is summarized in Table 2 according to market type, soil temperature and flower removal. About 25 more (p<0.05) shelled material was recovered on average for runner versus the spanish peanuts. Average recovery from ambient and heated soils was more (p<0.05) than 10 greater compared to chilled soils. A modest increase in recovery was observed for plots in which flowers had been removed, but was not statistically significant across the entire matrix. For these small plots, accurate yield predictions are not possible; however, this data demonstrates the established understandings that runner market types typically yield more than spanish market types, and at equivalent growing days coupled with full irrigation and optimal inputs, peanuts grown in warmer climates are expected to have greater yields.
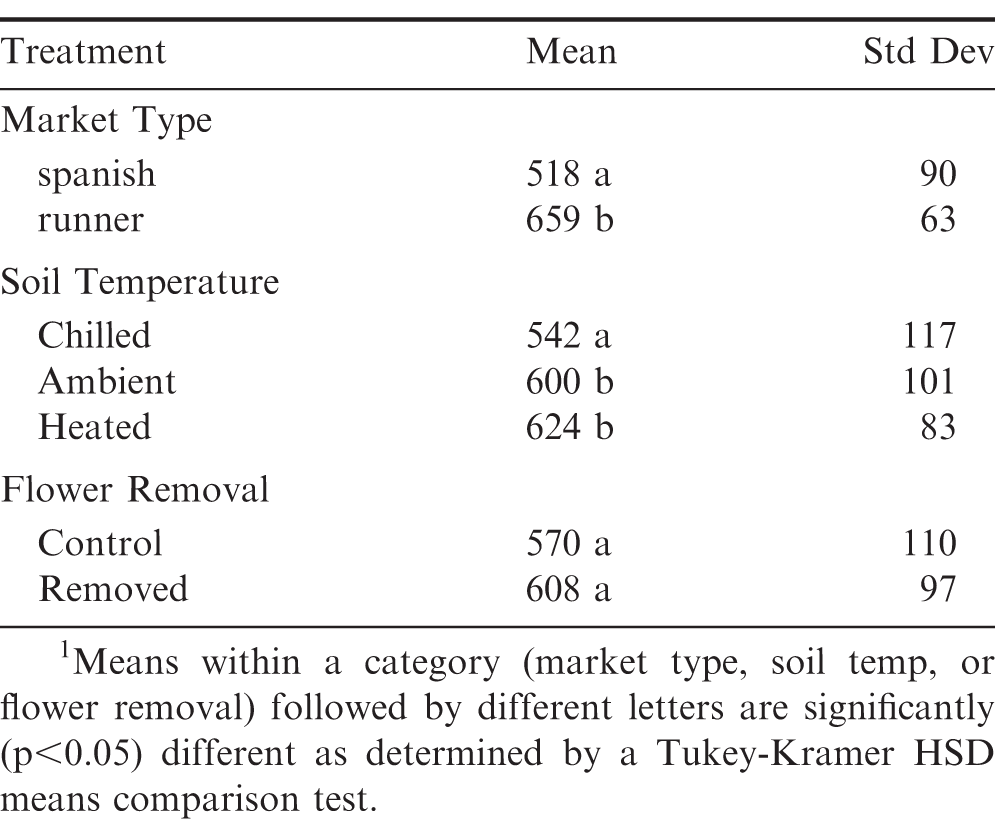
Mean1 and standard deviation of total shelled material (grams) recovered per m experimental row grouped by market type, soil temperature and late season flower termination.
The relative () of various size classifications after shelling are summarized in Figure 3. For runner peanuts, regardless of soil temperature or flower removal protocol, Jumbo's averaged about 50 of all shelled material, followed by Mediums at about 35, with the remaining approximate 15 comprising No1's, sound splits and oilstock. In the case of spanish peanuts, regardless of soil temperature or flower removal protocol, the No1 size comprised about 60 of shelled material, with Jumbo's comprising about 30 and the remaining approximate 10 comprised of oilstock and sound splits. It is emphasized that while similar nomenclature for runner and spanish sized kernels are used according to convention, i.e. Jumbo and No1, the actual sizes of these classes differ for the 2 market types as defined in the methods.
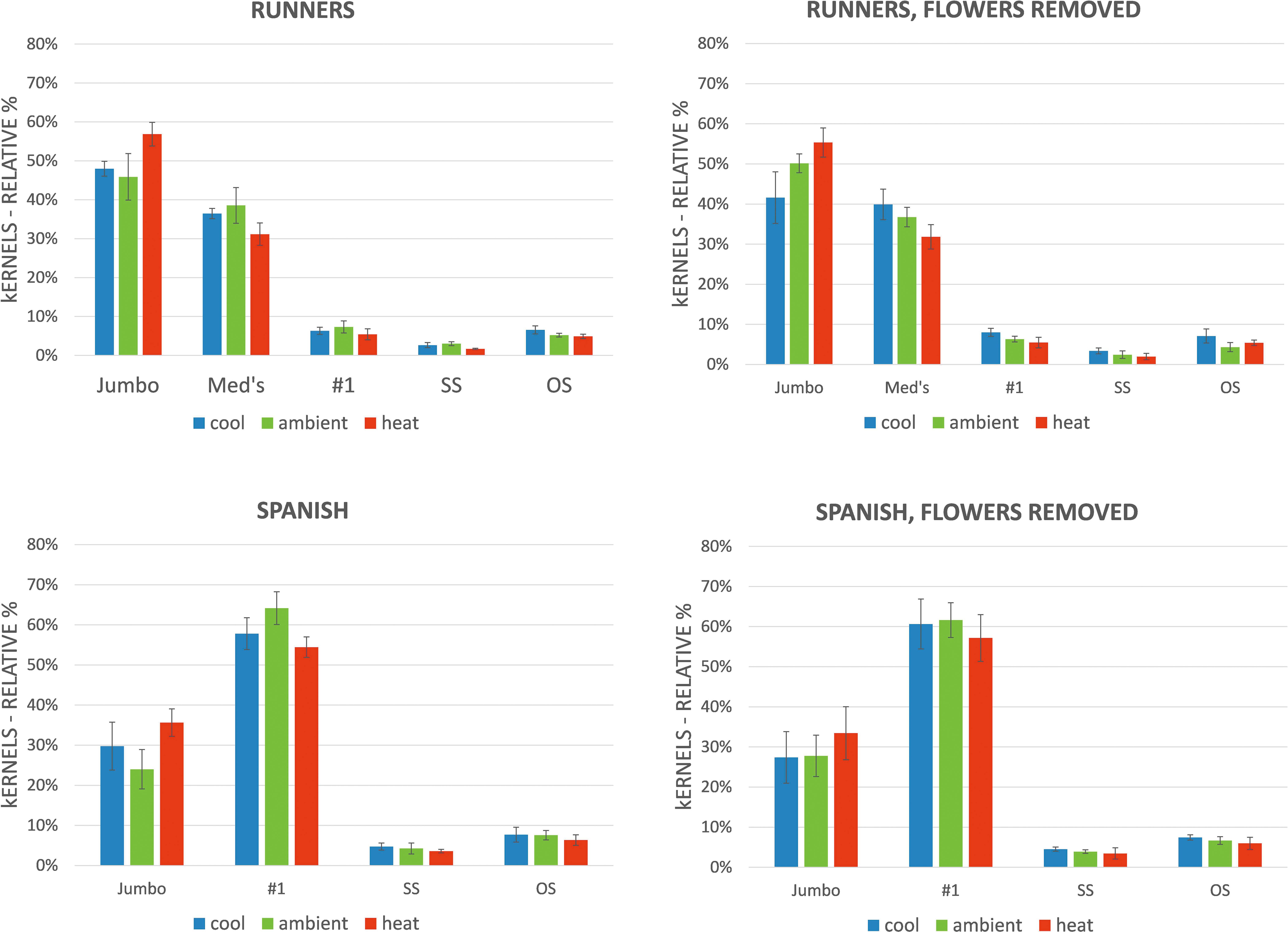
The relative () of various size classifications generated after shelling runner and spanish peanuts. Bars are averages of 4 replications and error bars are resulting standard deviations. Data are grouped according to market type, soil temperature and late season flower termination.
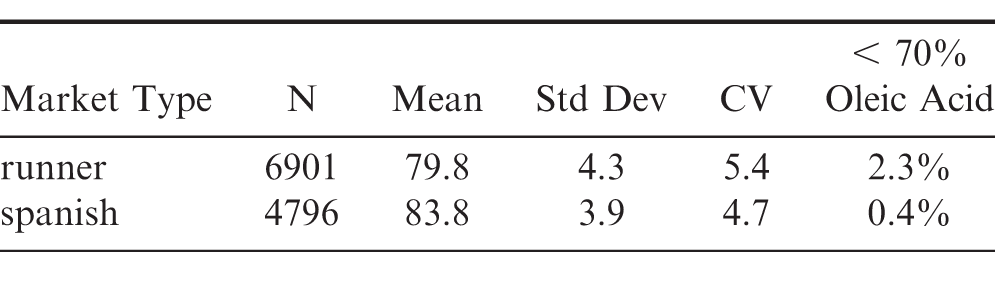
A total of 11,697 individual seeds were scanned for oleic acid () via NIR after harvesting, handling and shelling to simulate commercial practices. Summary statistics for the 6901 runner and 4796 spanish kernels are provided in Table 3. More runner peanuts were analysed as 3 size classes were prepared for runners: Jumbo's, Medium's and No1's per industry convention, versus only Jumbo's and No1's for the spanish. The overall mean was significantly (p<0.05) greater for spanish compared to the runner peanuts and the variation around the mean was less for the spanish kernels (Table 3). Shifts in average oleic acid () across the two populations are attributed to natural variation observed among high O/L peanut varieties. The decreased variation observed for the spanish peanuts likely primarily reflects the more determinate nature of the spanish market type as oleic acid () is a function of maturity and spanish varieties are expected to have a more consistent maturity profile at harvest.
Summary of oleic acid () for individual runner and spanish peanuts. At harvest, these samples were collected after simulating commercial processing as described in the manuscript.
Runner (Table 4) and spanish (Table 5) peanuts were further analyzed according to size, soil temperature and late season flower termination. For runners, increased kernel size significantly (p<0.05) affected average oleic acid, with Jumbo's having the greatest average oleic acid (), followed by Mediums and then No1's (Table 4). This is attributed to the documented correlation with increasing kernel size and increasing maturity (Pattee et al, 1982). A larger difference in average oleic acid () was observed between No1's and Mediums, compared to Medium's and Jumbo's. Among harvested runner peanuts, kernels from heated pod zones had significantly (p<0.05) greater mean oleic acid () compared to chilled and ambient soils (Table 4). This means that within a given size classification, there were still significant shifts in this key fatty acid chemistry depending on pod zone temperature. Such increases in oleic acid (and established, corresponding decreases in linoleic acid) are important, as these shifts continue to maximize shelf life. For spanish kernels, size (Jumbo's or No1's) did not affect mean oleic acid () whereas a significant increase was observed when comparing means of heated versus ambient or chilled soils (Table 5).
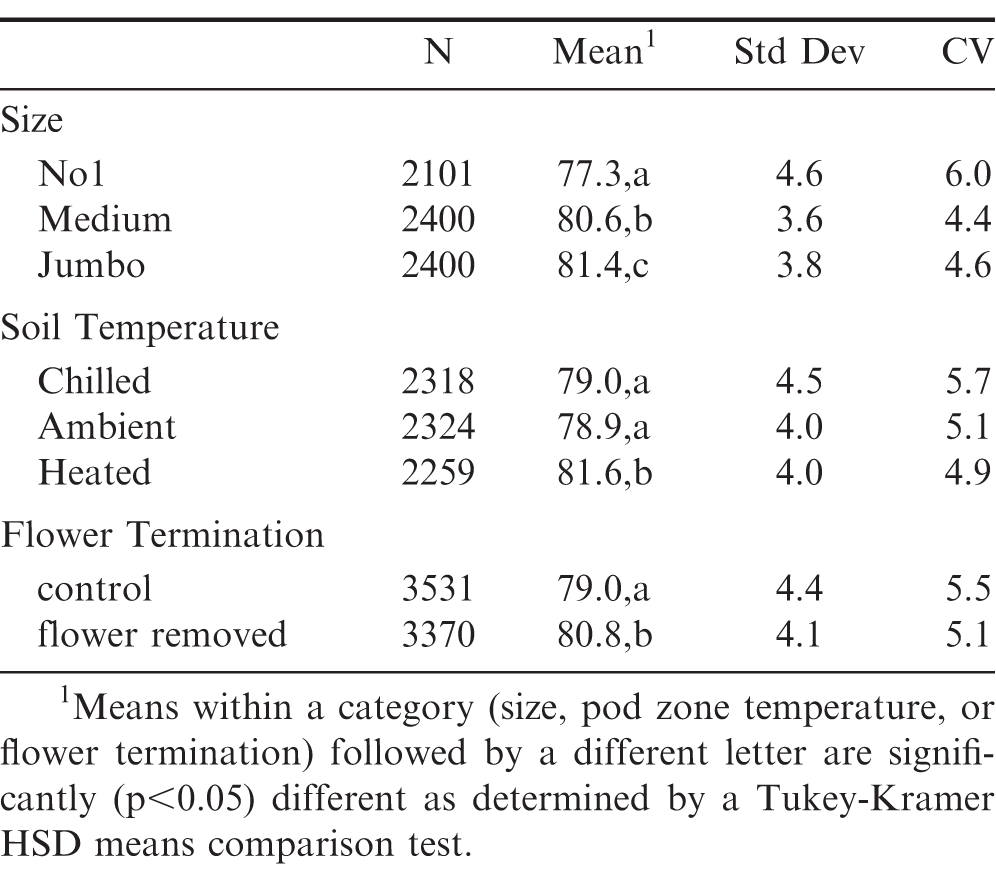
Summary statistics for oleic acid () of individual runner kernels as a function of size, pod zone temperature and late season flower termination.
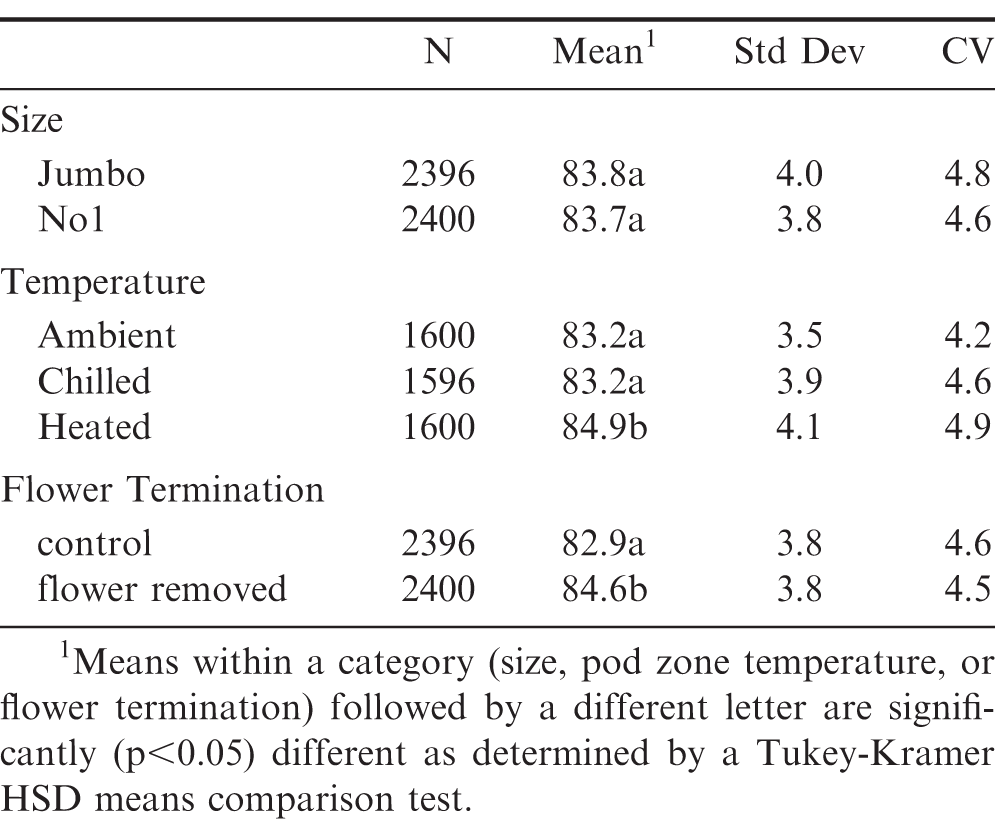
Summary statistics for oleic acid () of individual spanish kernels as a function of size, pod zone temperature and late season flower termination.
The increased oleic acid () of spanish kernels coupled with their decreased variation are ideal for many confectionary applications; however, the decreased yield of spanish varieties is an obvious economic disadvantage. Late season flower termination could be a strategy to not only improve farmers' stock yield and grade, but improve single kernel maturity distribution for runner peanuts. Late season flower termination increased (p<0.05) the overall mean compared to control runner peanuts (to a value closer to that of spanish) and decreased variation around this population (Table 4). For spanish kernels, late season flower termination also increased (p<0.05) the overall mean compared to control spanish peanuts and slightly decreased the coefficient of variation around this population (Table 5). Trends for mean oleic acid () for runner peanuts as a function of kernel size, soil temperature and late season flower termination are presented in Figure 4.
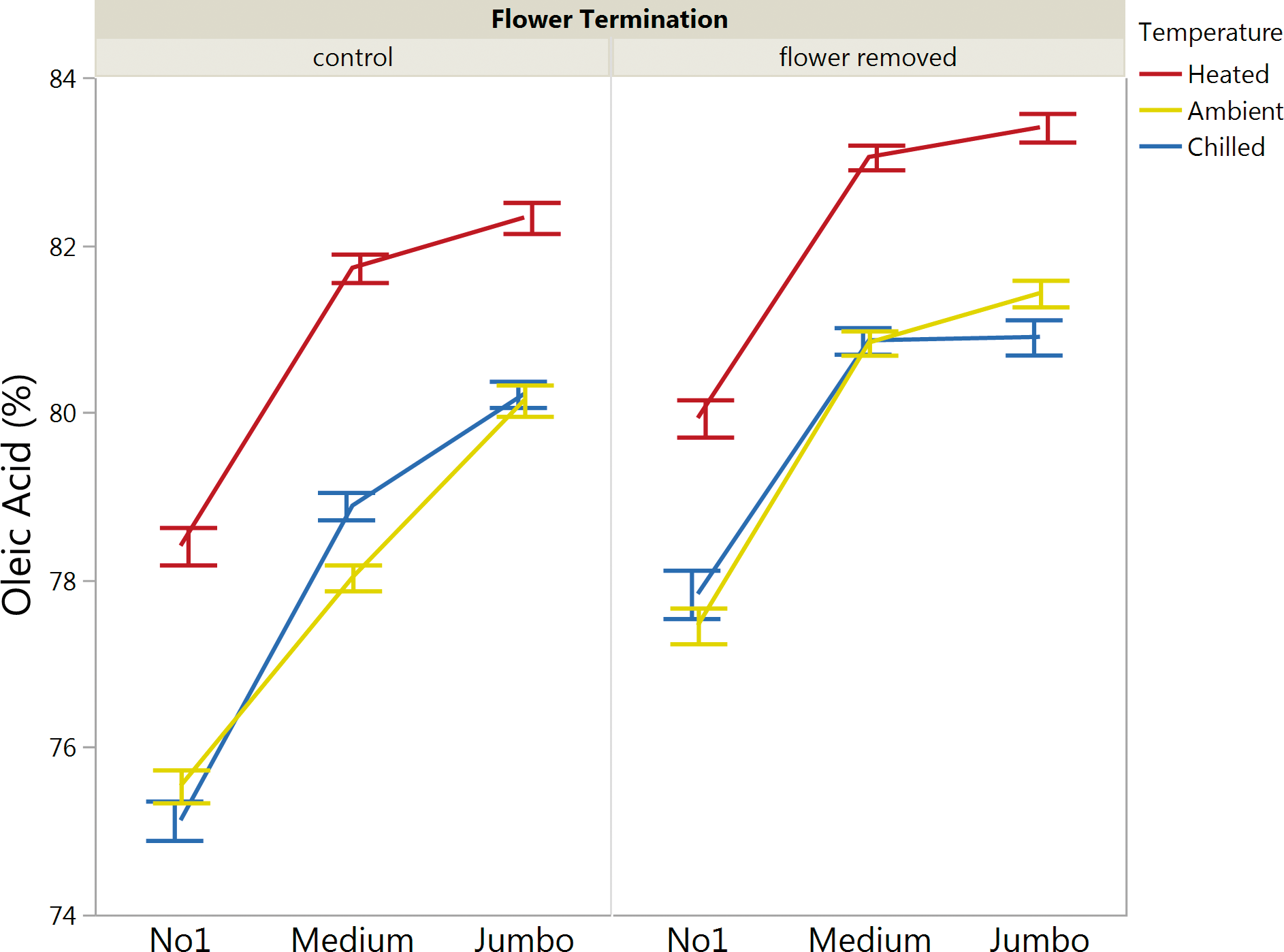
Average oleic acid () of individual runner kernels as a function of size, late season flower termination and soil temperature. Bars are means and error bars are standard error of means.
Flowers appearing later than approximately 90 days after planting (late season flowers) do not have enough time to make viable, mature peanuts given typical cultural practices. When this occurs, the plant is allocating photosynthetic resources into these flowers that could be used to mature the existing pods, hence the hypothesis that late season flower termination improves maturity distributions and single kernel oleic acid content on. Peanuts were also harvested and sorted according to mesocarp color into maturity classes (Williams and Drexler, 1981) for oleic acid () measurements. This data for runner peanuts (N 5402), averaged according to maturity class, soil temperature and late season flower termination is summarized in Figure 5. Maturity significantly (p<0.05) affected mean oleic acid () as did increasing soil temperature and late season flower termination, each of which increased oleic acid () across maturity samples (Figure 5).
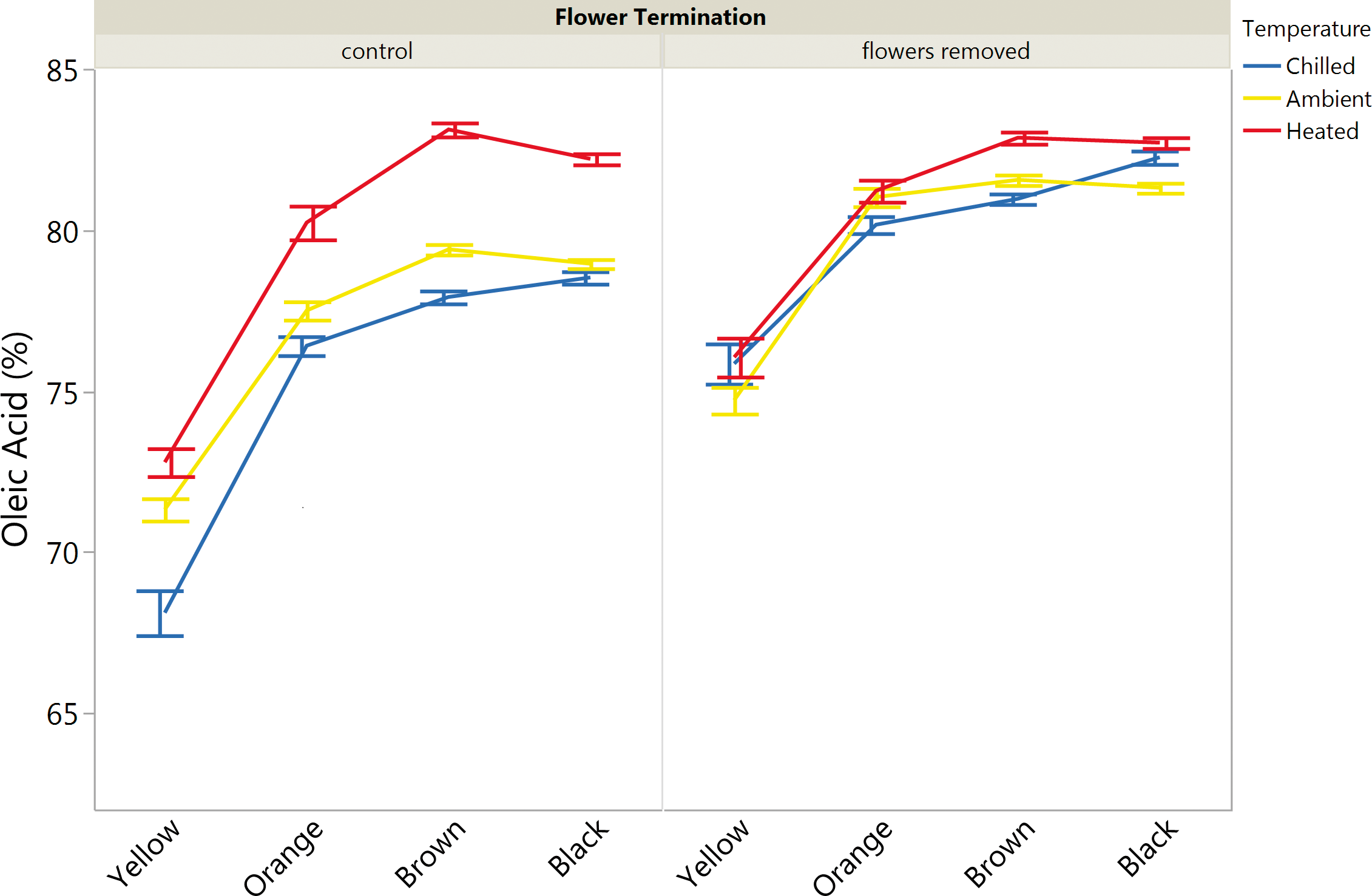
Average oleic acid () of individual runner peanuts as a function of maturity, late season flower termination and soil temperature. Bars are means and error bars are standard error of means.
A primary consideration in the industrial trade of high oleic lots is the percentage of kernels within a lot that do not meet the threshold fatty acid chemistry limits set for high oleic peanuts. While values may vary slightly, a typical threshold is an O/L ratio of 11.0 or greater (Sweigart et al, 2011). For these populations of spanish and runner high oleic peanuts experimentally processed to simulate commercial processing, approximately 2.3 of runner peanuts harvested had oleic acid values below 70, whereas spanish peanuts had only 0.4 of peanuts below 70 oleic acid (Table 3). Given the inherent and expected error in NIR measurement of oleic acid () and the typical observed separation of conventional and high oleic peanuts, peanuts with < 70 oleic acid measured in this study via NIR would very likely not meet the industry observed threshold for high oleic peanuts. Prior to planting, runner seed had about 3.2 peanuts below 70 oleic acid and spanish about 0.6 (Table 1), values that generally agree with these harvested progeny. As only peanuts with an oleic acid () greater than 75 measured by NIR were planted, i.e. all efforts were made to exclude non high oleic peanuts as contamination, it was hypothesized that all runner peanuts harvested at maturity would have oleic acid () values greater than 70, i.e. the purity of the progeny would be 100. Table 6 provides a breakdown of the runner peanuts not meeting threshold. When separating the peanuts by size, 87 of the peanuts not meeting the 70 oleic acid threshold were No1-sized peanuts. When comparing by soil temperature regime, 67 of the non-compliant peanuts were produced in the chilled soils. Late season flower termination reduced the peanuts not meeting the 70 oleic acid threshold by almost 50 to those harvested from the untreated control plots. These factors taken as a whole suggest maturity impacts on peanuts not meeting the minimal threshold for a field planted with a pure supply of high oleic peanuts, grown under ideal conditions, and harvested at maturity should be minimal, i.e. 2 or less, with the majority of runner peanuts not meeting threshold being expected in the No1 size class; however, further work is needed on a larger scale to verify these conclusions.
Distribution (N159) of runner peanuts less than 70 oleic acid for various categories.
For current experiments, late season flowers were hand removed; however, work over three years and multiple growing locations (small plot work) demonstrated that the input diflufenzopyr (BASF Biosciences) was effective at terminating late season flowers with corresponding approximate 400-500 lb/acre yield increases with irrigation while also improving farmer stock grade (Lamb et al., 2017). Research is actively ongoing to evaluate this technology at commercial levels on yield and grade, in addition to its impact on post-harvest quality, including single seed oleic acid () distributions.
Summary and Conclusions
Strong market type, soil temperature, size and maturity effects on oleic acid () among individual kernel populations of spanish and runner peanuts were observed. Implementation of a late season flower termination protocol strongly, and positively, influenced high oleic chemistry of runner peanuts. Data from this study provides an unprecedented understanding of expected natural variation in high oleic chemistry on the single kernel level
Acknowledgements
The authors gratefully acknowledge Mrs. C.M. Baker for conducting the NIR measurements. The authors also acknowledge the technical support staff at the National Peanut Research Laboratory including, Staci Ingram, Larry Powell, Hank Sheppard, Dan Todd and others for designing and installing the instrumentation and control systems, planting, managing, and harvesting the peanuts, then processing the samples after harvest.
Literature Cited
P.C., Andersen, K., Hill, D.W., Gorbet, B.V. Brodbeck, (1998). . Fatty acid and amino acid profiles of selected peanut cultivars and breeding lines. . J. Food Comp. Anal . 11: 100 111.
J.C., Braddock, C.A., Sims, S.F. O'Keefe, (1995). . Flavor and oxidative stability of roasted high oleic acid peanuts. . J. Food Sci . 60: 489 493.
C.L., Butts, E.J. Williamsand T.H. Sanders. (2002). . Algorithms for automated temperature controls to cure peanuts. . Postharvest Bio and Tech . 24: 309 316.
K.D., Chamberlin, N.A, Barkley, B.L., Tillman, J.W., Dillwith, R., Madden, M.E Payton, and R.S. Bennett, (2014). . A comparison of methods used to determine the oleic/linoleic acid ratio in cultivated peanut (Arachis hypogaea L.). . Agric. Sci , 5, 227 237.
Chen, C.Y., Wang, M.L., Lamb, M.C., Dang, P.M. 2014. Recovery and purification of spanish high oleate peanut AT-9899. 2014, . Proc. Am. Peanut Res. Educ. Soc. 46, 35.
J.I., Davidson, and F.P. McIntosh. (1973). . Development of a small laboratory sheller for determining peanut milling quality. . J. Am. Peanut Res. Educ. Assoc . 5: 95 108
J.P., Davis, D.S., Sweigart, K.M., Price, L.L., Dean, T.H. Sanders, (2013). . Refractive index and density measurements of peanut oil for determining oleic and linoleic acid contents. . J. Amer. Oil Chem. Soc . 90: 199 206.
C.M., Klevorn, K.W., Hendrix, T.H., Sanders, and L.L. Dean, (2016). . Differences in development of oleic and linoleic acid in high- and normal oleic virginia and runner-type peanuts. . Peanut Sci . 43, 12 23.
Knauft, D.A., Gorbet, D.W., Norden, A.J., inventors; University of Florida Research Foundation, Inc., assignee. 2000 May 16, 2000. Enhanced peanut products and plant lines. US Patent 6063984.
M.C., Lamb, R.B., Sorensen, C.L., Butts, P.M., Dang, C.Y. Chen, and R.S. Arias, (2017). . Chemical interruption of late season flowering to improve harvested peanut maturity. . Peanut Sci . 44: 60 65.
J.B., Mugendi, C.A., Sims, D.W., Gorbet, S.F. O'Keefe, (1998). . Flavor stability of high-oleic peanuts stored at low humidity. . J. Amer. Oil Chem. Soc . 75: 21 25.
H.E., Pattee, E.B., Johns, J.A., Singleton, T.H. Sanders, (1974). . Composition changes of peanut fruit parts during maturation. . Peanut Sci . 1: 57 62.
Pattee, H.E., Pearson, J.L., Young, C.T., Giesbrecht, F.G. 1982. Changes in roasted peanut flavor and other quality factors with seed size and storage time. J. Food Sci. 47, 455.
D.S., Sweigart, C.A., Homich, D.A., Stuart, (2011). . Rapid single kernel refractive index test that differentiates regular from high oleic peanuts. . Proc. Am. Peanut Res. Educ. Soc . 43, 27 28.
E.J. Williams, and J.S. Drexler, (1981). . A non-destructive method for determining peanut pod maturity. . Peanut Sci . 8, 134 141.
Notes
- Director of Technical Services, JLA International, Albany, GA USA, a subsidiary of IEH Laboratories, Lake Forest Park, WA 98155 and Adjunct Faculty, Dept. of Food, Bioprocessing & Nutrition Sciences, North Carolina State University, Raleigh, NC 27695 [^]
- Chairman of the Board, JLA International, Albany, GA USA, a subsidiary of IEH Laboratories, Lake Forest Park, WA 98155 [^]
- Dan S. Sweigart, The Hershey Company, Hershey, PA 17033 [^]
- Research Biochemist, Research Agricultural Engineer, Research Agronomist, and Supervisory Research Food Technologist, respectively, USDA, ARS, National Peanut Research Laboratory, Dawson, GA 39842 [^]
- Professor, Crop, Soil, and Environmental Sciences, Auburn University, AL 36849 [^] Corresponding Author Email: jackdavis@jlaglobal.com
Author Affiliations