

Introduction
As a botanically indeterminate plant, flowering and fruit initiation occurs in peanut over a long extended time period during the growing season. Thus a wide range of maturity in peanut fruit exists at any given harvest date. In the United States, runner-type peanuts for edible markets are separated according to kernel size into four marketing categories: Jumbos (ride 8.3-mm by 19-mm slotted screen), Mediums (ride 7.1-mm by 19-mm slotted screen), Number 1s (ride 6.3-mm by 19-mm slotted screen), and US splits (ride 6.7-mm round screen). Size and maturity relationships do exist but are not absolute and kernels of varying physiological maturity are consistently present within size categories and maturity variations within size categories are common. Distributions of maturity within commercial marketing sizes vary enough such that flavor, roast color, shelf-life, and other quality estimators may be affected in final roasted products (Sanders, et al., 1995; Sanders, 1989). These studies revealed that immature kernels roast darker and develop more "fruity fermented off-flavor" and less "roasted peanutty flavor" than mature peanuts within the same size category. A larger percentage of immature kernels at harvest led to a greater potential for negative quality impact during processing. Development of production and handling strategies that would produce a more consistent maturity distribution pattern would improve flavor consistency and shelf-life. The literature is limited on ways to obtain a more consistent maturity distribution but has focused instead on determining a highest yielding, best economical, harvest predicting, and non-destructive harvest date based system called the hull scrape maturity method (Williams and Drexler, 1981). In this method, a representative sample of fresh pods from a field to be harvested is processed to remove the exocarp by pod scraping, pod sand blasting, or pressure washing, which is more currently used, to reveal the peanut pod mesocarp color which ranges from white (immature) to black (mature) depending on maturity. This method separates colored pods into six morphological maturity classes to determine optimal digging date (Williams and Drexler, 1981). Research focusing on the physiological changes in peanut kernels (primarily oil content and composition) concluded that kernel development is related to stages in the hull scrape maturity method. For example, total oil as a percentage of dry weight increased significantly through class 5 (orange hull color) and then decreased in class 7 (black hull color). Oil color was proportionate to maturity class and generally decreased with increasing maturity (Sanders, 1989). Environmental and cultural practices (rainfed vs. irrigated production and location) did impact oil color. Free fatty acids decrease and O/L (oleic acid/linoleic acid) ratio increase as peanuts mature, each of which promotes improved shelf life and flavor potential, and progress through the maturity classes. The degree of changes in oil characteristics occurred substantially as peanuts entered class 5 (orange) which was defined as the beginning point of physiological maturity. Peanut quality factors and storability were influenced by the overall maturity level of the harvested peanuts (Sanders, 1989).
The current state of the literature focuses on determining peanut maturity for harvest date decisions as well as variations in maturity and the impact on post-harvest processing characteristics. Understanding of prescribed late season flower termination at the farmer and processor level is lacking. The original focus was to determine if late season flower termination to minimize immature kernels would result in improvements in the consistency and overall maturity level of peanuts thus improving quality factors of edible shelled stock peanut and this pursuit is continuing. However, the impact on yield, sound mature kernels plus sound splits (SMK+SS), other kernels (OK), and seed quality at the farm level will determine producer adoption. The objective of this research was to address the effect of prescribed late season flower termination through hand flower removal and chemical means on farmer stock yield, SMK+SS, OK, and seed germination.
Materials and Methods
Research was conducted in 2012 through 2014 at the USDA/ARS NPRL Bolton Irrigation Research Farm on a Ruston sandy loam (Fine-loamy, siliceous, semiactive, thermic Typic Paleudults) near Dawson, GA (31.790411°, -84.512228°). Irrigated and non-irrigated treatments were included. Irrigation in 2012 was done with shallow sub-surface drip (which has shown no yield difference from overhead sprinkler) and overhead sprinkler was used in 2013 and 2014 (Sorensen et al., 2010). Planting dates were 17 April 2012, 26 April 2013, and 24 April 2014. The peanut cultivar used was GA06G (Branch, 2007) which is the predominate cultivar in the Southeast. Each plot consisted of 6 rows established on a 0.91 m row spacing with a 21 seed/m seeding rate. Plots were sprayed with a CO2 pressurized sprayer delivering 140 L/ha at 172 kPa using 8002VS flat fan nozzles. Plot length was 9.1 m and the yield and quality evaluation samples were taken from the 2 middle rows of each plot allowing 4 border rows to minimize drift. Production management followed best management practices for fertility and weed, disease, and insect control. Experimental design was a randomized complete block design with a factorial treatment arrangement and 4 replications of each treatment. Irrigation was managed by IrrigatorPro® for Peanuts at each site (Davidson et al., 1998; Davidson et al., 2000). Producer level factors included farmer stock yield, grade factors, and seed germination. A 1.5 kg sample was analyzed by the Alabama Federal State Inspection Service (Dothan, AL) for determining SMK+SS, OK, and other grade factors. A whole kernel subset of the grade sample was obtained, treated with a commercial seed treatment fungicide, and sent to the Georgia Department of Agriculture Seed Lab in Tifton, GA for germination analysis. Processing attributes include shelling outturn distribution, kernel density by size category, and hull density. Manufacturing attributes will include roasted peanut flavor (0-10 scale), near-infrared reflectance analysis for fat, sugar, moisture, protein and nitrogen, and oil chemistry analysis for peroxide values and free fatty acids. Year was regarded as random with fixed treatments and the main effects and interactions will be tested using appropriate error terms. The treatments consisted of differing rates of two compounds: diflufenzopyr-Na (2-(1-[([3,5-difluorophenylamino] carbonyl)hydrazono]ethyl)-3- pyridinecarboxylic acid), BASF Biosciences) and glyphosate (N-(phosphonomethyl)glycine (41%)). Both were applied at different rates and timings. A non-treated control (NT-control) and a daily hand flower removal control (HFR-control) were included. HFR-control was initiated at 100 days after planting and continued daily through 115 days after planting. The application timings were 100 and 110 days after planting (DAP). Glyphosate was previously marketed in the late 1980's as a peanut growth regulator (Quotamaker 75WSP, Monsanto) to enhance peanut yields applied at low sub-lethal rates applied at first bloom (Beasley, 2001). Glyphosate was included in this study but applied late season to minimize onset of late season flowering. The rates for glyphosate were based on previous studies conducted in Georgia and North Carolina focused on the detrimental effect of glyphosate drift onto peanuts from application in adjacent glyphosate-tolerant crops (Grey and Prostko, 2010; Lassiter et al., 2007). Glyphosate rates were 56, 112, and 168 g ai/ha which were approximately 8, 17, and 25% of the registered use rate for glyphosate-tolerate field crops. In these studies, the application timings were at the 75, 90, or 105 days after planting (DAP) which approximated to R3, R4, or R5 growth stages (Grey and Prostko, 2010; Lassiter et al., 2007; Boote, 1982). Diflufenzopyr rates evaluated were 8, 17, and 25 g ai/ha as recommended by BASF in personal correspondence based on research to terminate late-season fruit development in cotton (Gossypium hirsutum L.) and soybean (Glycine max L.) (Bowe et al., 2000).
The spray application timings were intended to interrupt late-season flowering such that new onset flowers would not have sufficient time to produce marketable size yet immature peanuts prior to harvest. Thus, timing of treatments was later in plant development and differed from the aforementioned glyphosate drift studies. Two methods were compared to determine timing including the hull scrape maturity profile board and a fruit initiation method.
The current hull scrape maturity profile for runner-type peanuts is divided into six morphological maturity classes and provides estimated length of time required for peanuts to progress to the next stage (Williams and Drexler, 1981). These consist of:
-
1 - White (14-16 days progression to Yellow 1)
-
2 - Yellow 1 (10-14 days progression to Yellow 2)
-
3 - Yellow 2 (10-14 days progression to Orange)
-
4 - Orange (12-14 days progression to Brown)
-
5 - Brown (09-12 days progression to Black)
-
6 - Black (19-21 days progression to harvest).
The target timing of application would be to interrupt flowering (either through flower termination/abortion or through interruption of pollination) to minimize harvest of pods in the White, Yellow 1, and early Yellow 2 classes. If successful, this would concentrate the majority of harvested pods into the Orange, Brown, and Black classes which correspond to Class 4 which was defined as the beginning point of physiological kernel maturity. The application timings were determined on the current hull scrape maturity and the expected maturity date of the GA 06G cultivar.
Working backward through the expected progression days of each class and allowing 12 days for flower to initial in-soil peg development (pers. commun. with Auburn University peanut geneticist, Dr. Charles Chen), the application dates range from 92 to 108 days which averages to be 100 days after planting (DAP). This 100 DAP treatment timing also corresponds to determining the timing based on fruiting initiation date in Irrigator Pro for Peanuts and calculating forward across the profile board. Fruit initiation date in Irrigator Pro for Peanuts is defined as:
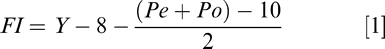
-
FI = Fruit Initiation Date,
-
Y = Date field was observed,
-
Pe = Average number of pegs per plant (defined as any visual growth of the gynophores), and
-
Po = Average number of pods per plant (Davidson et al., 1995).
Thus, adjustments in application due to delayed or accelerated fruit initiation date could be required depending on environmental factors between planting and fruit initiation. It was anticipated that a 20 day interruption in flower development at DAP 100 would be sufficient to prevent post-treatment flowers from developing into marketable sized prior to harvest as recommended by the hull scrape maturity method. Both methods were consistent each year in determining application timings which were approximately 100 or 110 DAP which corresponds to R5 and R6 growth stages, respectively (Boote, 1982).
Flower counts were initiated on the first application date and conducted daily in each treatment in a 1 m of row in each plot in block 2 of the randomized block design beginning at DAP 100 and continuing through DAP 115. Counts included both controls and all treatments but were limited to only one block within the randomized complete block design due to time and labor constraints. Although flower counts were limited to a single replication of each treatment, the HFR-control was conducted in all replications to allow statistical comparisons with the NT-control and chemical treatments. The flower counts provide data on flower addition rates over time for the chemical treatments versus the NT-control and the HFR-control. Significance was tested using the pdiff option in LSmeans statement of PROC GLIMMIX.
Results and Discussion
Irrigation requirements in 2013 were substantially lower than in 2012 and 2014 due to adequate and consistent rainfall (Figure 1). Rainfall in 2013 totaled 66.7 cm during the production season and irrigation applied was 11.4 cm. Conversely in 2012 and 2014 rainfall during the growing season totaled 34.4 cm and 26.5 cm with irrigation applied totaling 30.8 and 37.8 cm, respectively. The drought in 2014 was more severe late-season negatively impacting pod fill (Figure 1). Significant damage from lesser cornstalk borer (Elasmopalpus lignosellus L.) resulted in the 2014 non-irrigated plots even though chlorpyrifos at 2.2 kg/ha was applied (Lorsban 15G insecticide Dow AgroSciences, Indianapolis, IN).
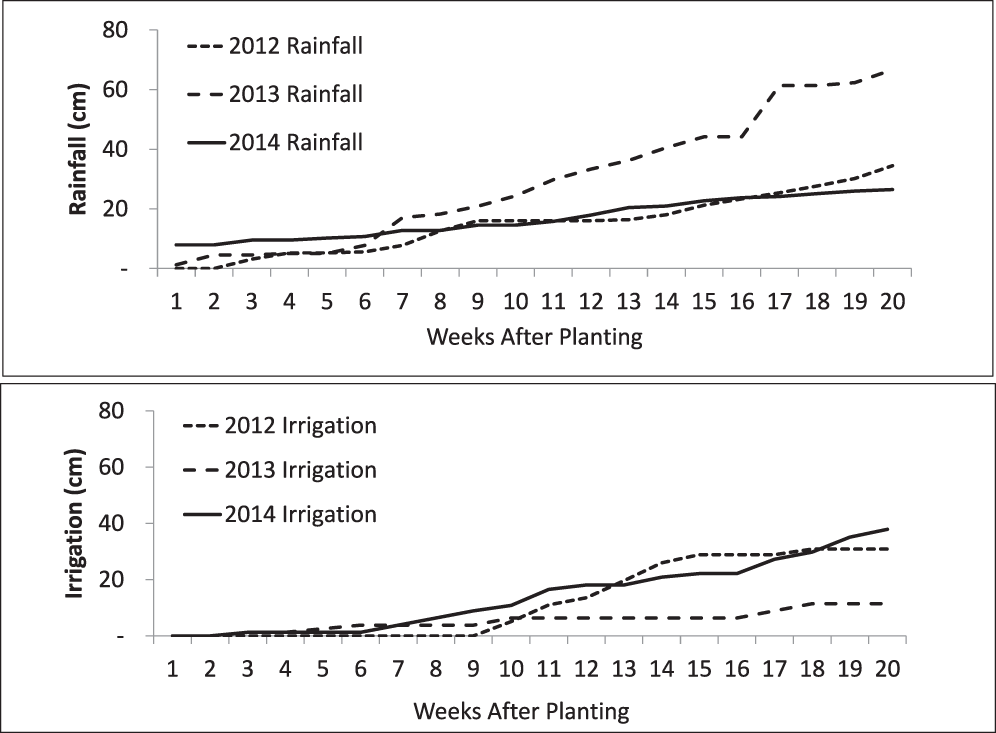
Cumulative weekly rainfall and irrigation distributions for the 2012-2014 cropping seasons at the USDA/ARS Bolton Irrigation Research Farm, Dawson, GA.
Daily flower counts were initiated on the date that the treatments were applied and continued for 15 days. Counts were conducted in same 1 m of row section of each treatment in Block 2 at the same time each day (6:30 am EST). Since time and labor constraints prevented flower counts in all replications of each treatment, no statistical inference is made. Counts in the HFR-control were conducted immediately before flowers were removed. The flower counts illustrate differences in the efficacy among the treatment groups (Figure 2). The data are graphically presented by treatment group for the cumulative flower counts irrigated and non-irrigated plots averaged over the 2012-2014 cropping seasons. Cumulative counts are presented to better show trends as opposed to flower counts per day which have more variation. In the non-irrigated treatments, the counts on 1 day after treatment (DAT) were tightly grouped at approximately 10 flowers per 1 m of row (Figure 2). When combined over rates and timings, by 15 DAT the non-irrigated cumulative flower counts were 240, 210, 90, and 118 which equated to a daily flower addition rate over the 15 day period of 16, 14, 6, and 8 for the NT-control, HFR-control, glyphosate, and diflufenzopyr treatments, respectively. On a percentage basis HFR-control, glyphosate, and diflufenzopyr resulted in 14, 166, and 104 percent lower flower counts in the non-irrigated plots than the NT-control (Figure 2).
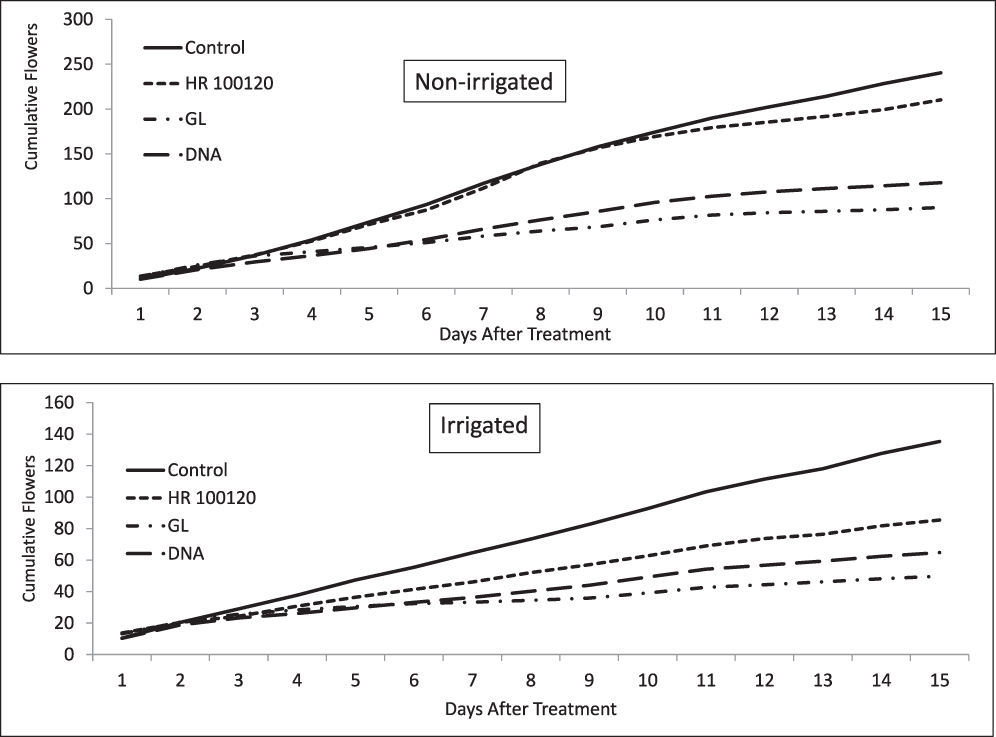
Cumulative flowers counts per 1 m of row in Non-irrigated and Irrigated peanuts at the USDA/ARS Bolton Irrigation Research Farm, Dawson, GA (2012-2014).
Flower counts in the irrigated treatments had less flowering activity but similar trends compared to the non-irrigated counts. Cumulative counts (combined over rates and timings) over the 15 DAT period were 135, 85, 49, and 64 equating to a daily addition rate of 9, 6, 3, and 4 for the NT-control, HFR-control, glyphosate, and diflufenzopyr treatments, respectively. The percentage change in the irrigated plots for HFR-control, glyphosate, and diflufenzopyr were 59, 166, and 104 percent lower than the NT-control (Figure 2). The chemical treatments were effective in minimizing late season flowering compared to both the NT-control and HFR-control treatments.
The main effect of irrigation was significant for yield, SMK+SS, and OK: yield (6,515 kg/ha irrigated vs. 3,309 kg/ha non-irrigated: P0.0001), SMK+SS (73.8% irrigated vs. 70.2% non-irrigated: P0.0001), OK (3.7% irrigated vs. 4.7% non-irrigated: P0.001) and germination (84.3% irrigated vs. 76.3 non-irrigated: P0.001). The main effect of year for yield, SMK+SS, and OK was also significant at P0.0001. Application timing was not significant and was removed from the statistical model to achieve greater replication. However, the interaction of irrigation by treatment and rate was not significant and thus are presented separately for irrigation, treatment, and rate.
Irrigated and Non-irrigated Peanut Yield
Glyphosate did not affect irrigated or non-irrigated yield compared to the NT-control and HFR-control. Although glyphosate rates were not significant, there was an inverse trend of rate in both irrigated and non-irrigated peanut yield consistent with previous research in Georgia and North Carolina. These trends are important when considering rate risks as glyphosate rates greater than 160 g/ha were shown to result in a significant yield reduction (Grey and Prostko, 2010; Lassiter et al., 2007). Diflufenzopyr at the 17 and 25 g/ha rate increased peanut yield by 446 and 567 kg/ha, respectively compared to the NT-control. Diflufenzopyr at the 17 and 25 g/ha rates also increased yield compared to the HFR-control and glyphosate at the 112 and 168 g/ha rates, respectively (Table 1). At the current loan rate for runner-type peanuts, the increased yield at the 17 and 25 g/ha diflufenzopyr rates provides $146.51 and $186.31 increased revenue per hectare compared to the NT-control. Diflufenzopyr is not currently available commercially but instead is used as a co-product in existing commercially labelled herbicides. Thus a cost per hectare for diflufenzopyr is not available for economic analyses. Non-irrigated yield did not differ among the treatment groups and rates (Table 1).
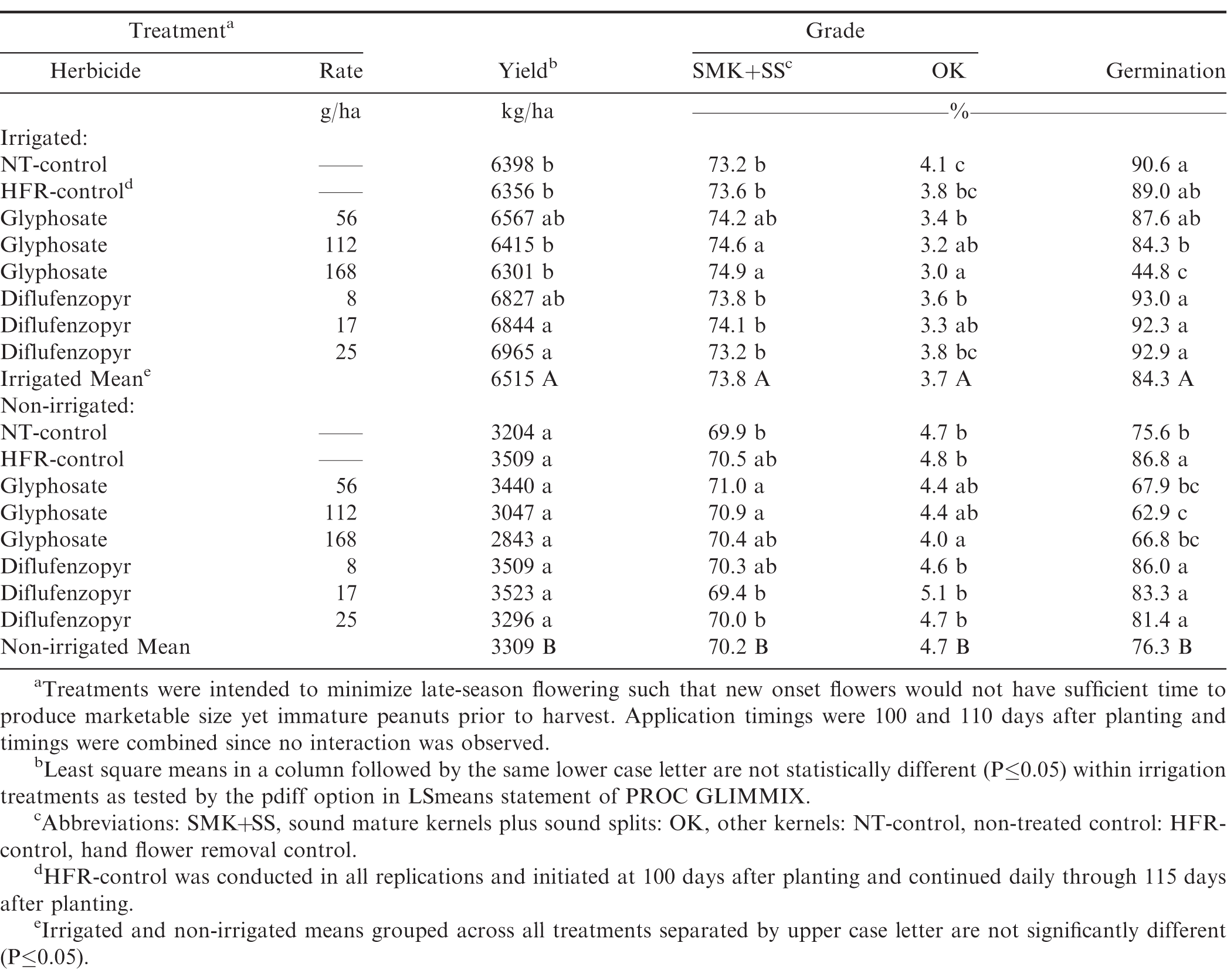
Peanut yield, grade, and seed germination as influenced by selected, late-season treatments, including sub-lethal glyphosate, applied with the intent of disrupting/stopping late season flowering; irrigated and non-irrigated production; 3-yr data pool: 2012-2014 crop years.
Irrigated and Non-irrigated SMK+SS and OK
Glyphosate at the 112 and 168 g/ha rate increased irrigated SMK+SS over all of the controls and treatment groups. Irrigated OK were reduced at all glyphosate rates compared to the NT-control. OK reductions were also observed at the 8 and 17 g/ha rates of diflufenzopyr compared to the NT-control (Table 1). In the non-irrigated treatments, glyphosate improved SMK+SS at the 56 and 112 g/ha rates and non-irrigated OK were reduced at the 168 g/ha rate compared to the NT-control. However, the increased value per ton associated with increased SMK+SS and decreased OK for either chemical treatment are not sufficient, to cover the costs of glyphosate and application. The improvements in both SMK+SS and OK in the glyphosate and diflufenzopyr groups are attributed to the efficacy in flower termination. Advanced maturity, as determined by the hull scrape maturity method, was consistently observed in the glyphosate and diflufenzopyr treatments compared to the controls (data not shown). Benefits could accrue to the post-harvest processing sectors as improvements in maturity distributions contribute to improved processing characteristics including peanut flavor, color, and shelf-life and reduce the risk for negative impacts during processing (Sanders, at el., 1995; Sanders, 1989). Impacts beyond the farmer stock level are forthcoming.
Irrigated and Non-irrigated Germination by rate
Glyphosate reduced irrigated seed germination at the 112 g/ha rate compared to the NT-control and all diflufenzopyr treatments and the 168 g/ha glyphosate rate reduced germination compared to all controls and treatments (Table 1). A reduction in irrigated germination (3%: not significant) at 56 g/ha rate was also observed. Glyphosate at the 112 g/ha rate reduced non-irrigated germination compared to the NT-control and HFR-control (Table 1). Reduced germination from glyphosate has been demonstrated in studies in other crops (Roider, et al., 2007; Steadman et al., 2006; Yasour et al., 2007; Yenish and Young, 2000). Others studies showed no impact of glyphosate on germination (Duke et al., 2003; Grey and Prostko, 2010). The trend of negative seed quality associated with the increasing rates of glyphosate indicates that seed supplies should not be derived from peanuts where glyphosate was applied, even if unintentional, during the reproductive and maturation stages. Diflufenzopyr showed numeric but not statistically different improvements in irrigated germination compared to the NT-control or HFR-control but increased irrigated and germination over the higher glyphosate rates. Diflufenzopyr at all rates increased non-irrigated germination over the NT-control and glyphosate treatments (Table 1).
In summary, this three-year study focused on the impact of late-season flower termination on peanut yield, SMK+SS, OK, and seed germination produced in non-irrigated and irrigated regimes. Two chemical treatments at very low, sub-lethal rates (Glyphosate and Diflufenzopyr) were evaluated against two controls (NT-control and HFR-control). The rates of the chemical treatments were intended to interrupt late-season flowering for a short period without causing significant plant injury allowing continued maturation of existing pods. Non-irrigated yield was not affected by treatment group or rates. Diflufenzopyr increased irrigated yield at the 17 and 25 g/ha rate by 446 and 567 kg/ha compared to the NT-control which equates to increased gross revenue of $146.52 and $186.3 per hectare, respectively. Glyphosate did not affect peanut yield but did provide improvements in SMK+SS and OK. However, the reductions in seed quality clearly show that late-season applications of glyphosate should be avoided at any rate especially for fields designated for seed production. Diflufenzopyr showed improvements in seed quality in non-irrigated peanuts.
Literature Cited
Beasley, J.P 2001 Roundup and Peanuts In Peanut Pointers Tifton, GA: University of Georgia Cooperative Extension Service Newsletter. Vol. 38 No. 7.
K.J Boote, (1982). Growth stages of peanut (Arachis hypogea L.). Peanut Sci 9: 35- 40.
Bowe, S.J, Rademacher, W., Newson, L.J., and Ferguson, G.P 2000 Use of semicarbazone plant growth regulator for crop yield enhancements. U.S Patent: US6150302 A. Nov. 21, 2000.
W.D Branch, (2007). Registration of 'Georgia-06G'peanut. J. Plant Reg 1: 120- 120.
J.I., Davidson, C.T Bennett, T.W Tyson, J.A Baldwin, J.P Beasely, M.J Bader, and A.W Tyson (1998). Peanut irrigation management using Exnut and Moisnut computer programs. Peanut Sci 25: 103- 10.
Davidson, J.I., Jr., M.C Lamb, C.L Butts, E.J Williams, and M Singletary 2005 Applications of expert systems in peanut production In: H.E Patteeand H.T Stalker, (eds.) Advances in Peanut Science Amer. Peanut Res. Educ. Soc. Inc., Stillwater, OK.
Davidson, J.I., Jr., M.C Lamb, and D.A Sternitzke 2000 Farm Suite - Irrigator Pro (Peanut Irrigation Software and Users Guide), The Peanut Foundation: Alexandria, VA.
S.O., Duke, A.M Rimando, P.F Pace, K.N Reddy, and R.J Smedaj (2003). Isoflavone, glyphosate, and aminomethylphosponic acid levels in seed of glyphosate-treated, glyphosate-resistant soybean. Agric. Food Chem 51: 340- 344.
T.L., Grey, and E.P Prostko (2010). Physiological effects of late season glyphosate applications on peanut (Arachis hypogaea L.) seed development and germination. Peanut Sci 37: 124- 28.
B.R., Lassiter, I.C Burke, W.E Thomas, W.E Pline-Srnic, D.L Jordan, J.W Wilcut, and G.G Wilkerson (2007). Yield and physiological response of peanut to glyphosate drift. Weed Technol 21: 954- 60.
C.A., Roider, C.A Jones, S.A Harrison, and J.L Griffin (2007). Wheat response to simulated glyphosate drift. Weed Technol 21: 1010- 1015.
T.M Sanders, (1989). Maturity distribution in commercially sized Florunner peanuts. Peanut Sci 2: 91- 95.
Sanders, T.M., H.E Pattee, J.E Vercellotti, and K.L Bett 2005 Advances in peanut flavor quality In: H.E Patteeand H.T Stalker, (eds.) Advances in Peanut Science Amer. Peanut Res. Educ. Soc. Inc., Stillwater, OK.
Sorensen, R.B., Nuti, R.C., Lamb, M.C 2010 Yield and economics of shallow subsurface drip irrigation (S3DI) and furrow diking Crop Management DOI: 10.1094CM-2010-1220-01-RS.
K.J., Steadman, D.M Eaton, J.A Plummer, D.G Ferris, and S.B Powles (2006). Late-season non-selective herbicide application reduces Lolium rigidum seed numbers, seed viability, and seedling fitness. Australian J. Agri. Res 57: 133- 141.
E.J., Williams, and J.S Drexler (1981). A non-destructive method for determining peanut pod maturity. Peanut Sci 8: 134- 41.
H., Yasour, J Riov, and B Rubin (2007). Glyphosate-induced male sterility in glyphosate-resistant cotton (Gossypium hirsutum L.) is associated with inhibition of anther dehiscence and reduced pollen viability. Crop Prot 26: 363- 369.
J.P Yenish, and F.L Young (2000). Effect of preharvest glyphosate application on seed and seedling quality of spring wheat (Triticum aestivum). Weed. Technol 14: 212- 217.
Notes
- Research Food Technologist, Research Agronomist, Research Agricultural Engineer, Research Biochemist, Research Pathologist. USDA, ARS, National Peanut Research Laboratory, 1011 Forrester Drive, S.E., Dawson, GA 39842; 2Professor. Crop, Soil, and Environmental Sciences, Auburn University, Auburn, AL 36849 [^]
- Professor. Crop, Soil, and Environmental Sciences, Auburn University, Auburn, AL 36849 [^] *Corresponding author's E-mail: Marshall.Lamb@ars.usda.gov
Author Affiliations