

Introduction
Peanut ( Arachis hypogaea L.) growers rely on preemergence and postemergence herbicides for managing weeds, but weed escapes are common due to environmental conditions, application errors, and more recently the evolution of herbicide-resistant weeds (Heap, 2014). Weeds that survive control measures can compete with peanut and cause up to 60% yield loss depending on weed species and population densities (Barbour and Bridges, 1995). Furthermore, they could produce seed perpetuating or even increasing weed seed banks.
It has been proposed that crops can be improved to be more competitive by increasing their ability to suppress weed growth and reproduction (Andrew et al., 2015; Jannink et al., 2000; McDonald, 2003). Similarly, several researchers have suggested that the ability of the crop to tolerate weed interference can be increased through breeding (Andrew et al., 2015). Weed suppressive ability and tolerance to interference are two distinct traits that might operate through similar processes. The former is the ability of a crop to reduce weed growth, and the latter is the ability of the crop to minimize yield loss when resources are limited due to weed interference (Andrew et al., 2015; McDonald, 2003). Weed suppressive ability is predominantly influenced by plant architectures that favor shading and rapid canopy closure (Andrew et al., 2015; Barbour and Bridges, 1995; Légère and Schreiber, 1989), and so is directly related to vegetative growth. Conversely, weed interference tolerance depends on the tradeoff between vegetative and reproductive growth (Andrew et al., 2015; McDonald, 2003) and can be a trait more difficult to characterize than weed suppressive ability (Watson et al., 2006). Thus, individuals that can redistribute photoassimilates to maintain grain or fruit production at the expense of vegetative growth would exhibit increased tolerance to interference because yield would not be reduced compared to individuals that favor vegetative growth. Interestingly, it is possible that individuals, which favor vegetative growth during interference might have higher weed suppression ability than individuals that favor reproductive growth. However, the former may be less tolerant to weed interference than the latter.
Peanut-weed interference has been studied to characterize potential yield loss, to determine optimum planting arrangements, and the critical period of competition when weed control actions are most needed (Agostinho et al., 2006; Hauser et al., 1975; Hauser and Buchanan, 1981; Place et al., 2010). Place et al. (2012) compared the response to weed interference of eight Virginia market type genotypes without finding clear differences among genotypes. However, all genotypes suffered significant yield losses resulting from weed interference. With the exception of the Place et al. (2012) study, which focused on peanut biomass and yield responses, there is no information about which morphological characteristics and changes in canopy architecture favor peanut weed suppressive ability and the variability among peanut cultivars to tolerate weed interference. The objectives of the present study were: 1) to determine whether peanut cultivars with different canopy architecture and growth habit differ in their weed suppressive ability and weed interference tolerance, and 2) to characterize morphological and physiological responses to differing levels of weed interference.
Materials and Methods
A field experiment was conducted at the West Florida Research and Education Center in Jay, FL during 2013, 2014, and 2015. The experimental site was a field that had been kept with a dense weed seed bank for the last 30 years. This field was selected to ensure high weed pressure in all plots and avoid issues related to patchiness common in fields with low weed seed banks. The weed community was comprised mainly of the dicotyledonous species sicklepod [Senna obtusifolia (L.) H.S. Irwin & Barneby], pitted morningglory ( Ipomoea lacunosa L.), smallflower morningglory [Jacquemontia tamnifolia (L.) Griseb.]; and the monocotyledonous species benghal dayflower ( Commelina benghalensis L.), goosegrass [ Eleusine indica (L.) Gaertn.], browntop millet [Urochloa ramosa (L.) T.Q. Nguyen], large crabgrass [ Digitaria sanguinalis (L.) Scop.], and crowfoot grass [ Dactyloctenium aegyptium (L.) Willd]. The soil was a Red Bay sandy loam (Fine-loamy, kaolinitic, thermic Rhodic Kandiudult) with pH 6.2 and 69% sand, 16% silt, and 15% clay. Every year the experiment was moved to an adjacent fallow area within the same field to avoid confounding residual effects from the previous year.
The treatments were three peanut cultivars and one advanced breeding line with different growth habit and canopy architecture and three weed interference conditions. The cultivars were ‘Bailey’ (erect growth and tall canopy height; Isleib et al., 2011), ‘Georgia-06G’ (semi-bunch, intermediate height; Branch, 2007), and ‘TUFRunner 727’ (prostrate growth and intermediate height; Gorbet and Tillman, 2013); the advanced breeding line ‘UFT312’ has very prostrate growth and short canopy height (Tillman, unpublished results). The weed interference conditions were no interference (weed-free control), early season interference, and full-season interference. These conditions were achieved by controlling weeds at different times during the growing season with a combination of herbicides and hand weeding. Additionally, we included a no-peanut control for all interference conditions to quantify weed growth potential in the absence of the crop, which allowed us to determine weed suppression by each cultivar. The experiment was arranged as a randomized complete block split-plot design with four replications.
Each season, fields were fertilized according to soil test recommendations, moldboard plowed, and beds were formed. All cultivars were planted in single rows at 20 seeds/m of row. In 2013, row spacing was 76 cm, and 91 cm in 2014 and 2015. Plots were four rows wide and 7.6 m long. Disease and insect management practices were implemented based on local standard recommendations.
All plots were treated with pendimethalin (Prowl H2O®, BASF Corporation, Research Triangle Park, NC) at 1,120 g ai/ha, which was incorporated with a field cultivator before planting. Similarly, all plots were treated with paraquat (Gramoxone Inteon, Syngenta, Greensboro, NC) at 140 g ai/ha plus a non-ionic surfactant at 0.25 % (v/v) (Induce, Helena Chemical Company, Collierville, TN) at cracking to eliminate weeds that emerged before peanut. This was done to avoid excessive weed interference that, based on previous studies, would have completely outcompeted the peanut preventing our ability to measure yield loss responses at levels commonly faced by growers. The full-season interference treatment did not have any additional weed control action after paraquat application. The no interference treatment had an application of a tank-mixture with imazapic (Cadre, BASF Corporation, Research Triangle Park, NC) at 70 g ai/ha, fluazifop-P-butyl (Fusilade DX, Syngenta, Greensboro, NC) at 420 g ai/ha, and non-ionic surfactant at 0.25 % (v/v) 4 and 8 wk after planting (WAP). The early season interference treatment had an application of the aforementioned tank-mixture only 8 WAP. Additionally, the no interference and early season interference treatments were hand-weeded as needed to ensure weed-free plots.
Peanut canopy height and width were determined at 4, 6, 8, 10, and 12 WAP by measuring five plants randomly selected from the two center rows of each plot. Four weeks before harvest, weed biomass was determined by harvesting all weeds (aboveground tissue) in a 1-m2 frame randomly located within each plot, and then drying the tissue at 65 C for 7 d. At maturity, two peanut plants per plot were harvested (including underground tissue, and number of leaves (i.e. trifoliates), total leaf area, and plant dry weight were determined. Before harvest, weedy plots were mowed above the peanut canopy and hand weeded if needed to avoid pod loss during harvest (Place et al., 2012). The two middle rows of each plot were dug with a conventional digger-shaker-inverter and were allowed to air-dry in the field for 5 to 7 d. Peanut was then picked with a two-row peanut combine and yield was determined at 10% moisture. Optimal peanut harvest time was determined using the hull scrape method (Williams and Drexler, 1981).
Main plots were cultivar, and weed interference durations were randomized as subplots. Data were analyzed with ANOVA using the Mixed Procedure of SAS (9.2 SAS® Institute Inc. Cary, NC 27513) considering peanut treatment, interference condition, year and their interactions as main effects (α=0.05). For peanut height and canopy width data, a repeated measurements analysis was conducted. A split-plot analysis was conducted for the remaining dependent variables. The Tukey-Kramer Honestly Significant Difference method was used for means separation tests (α=0.05).
Results and Discussion
Results were analyzed by year because of significant interactions (P<0.03) between year, cultivar, and interference treatments. Unusually intense rainfall events were received during the planting and establishment period between mid-April and late May 2014 (Figure 1). The UFT312 seed used in 2014 had lower vigor than the other two years, and crop stand was considerably lower (less than half) than the other cultivars. For this reason, UFT312 data were excluded from analyses in 2014. Within years, few interactions were observed between cultivar and interference (Table 1).
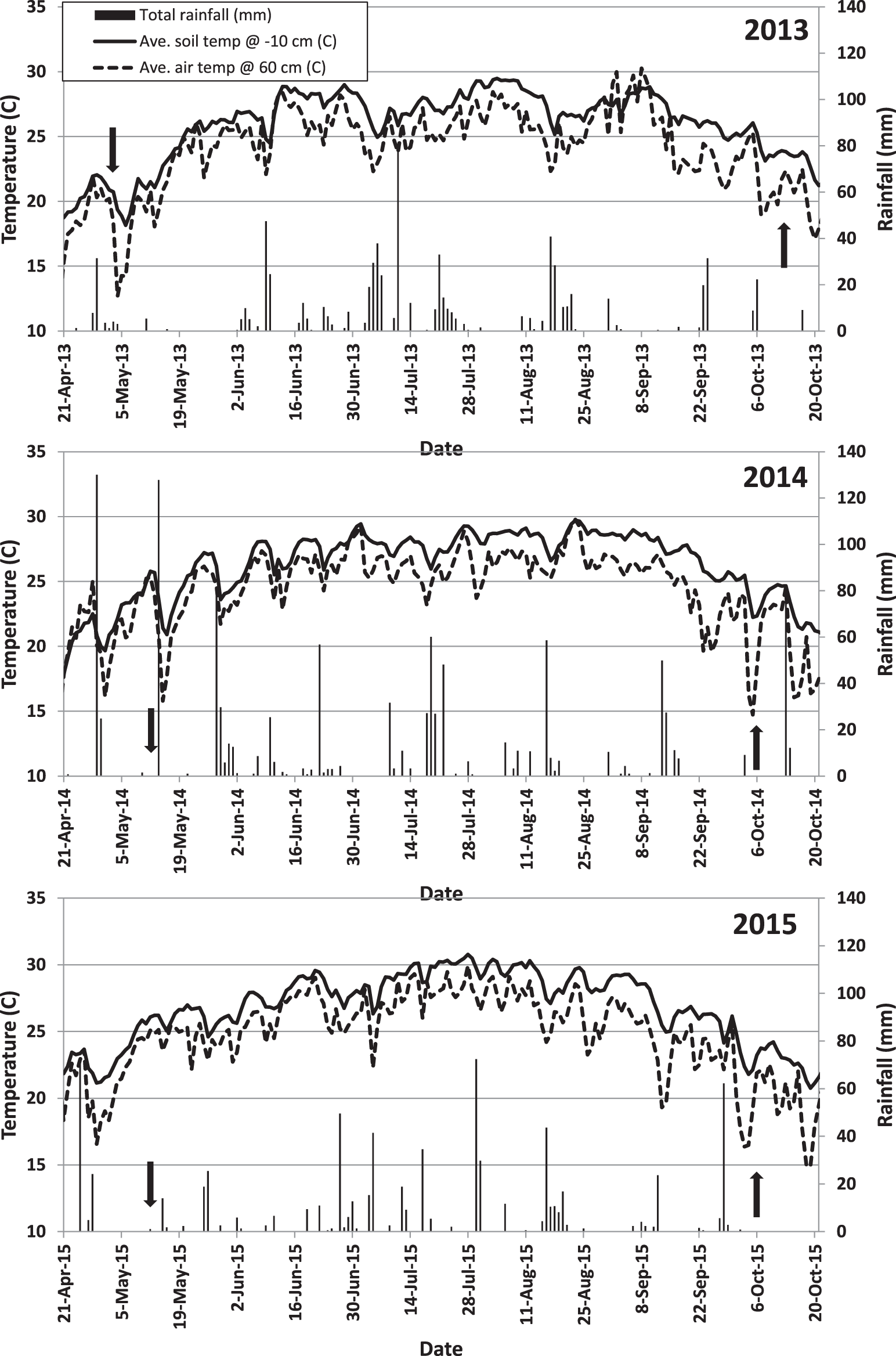
Daily air and soil temperatures and rainfall data at the research site in Jay, FL over the three year study. Downward and upward arrows indicate planting and harvest date, respectively.
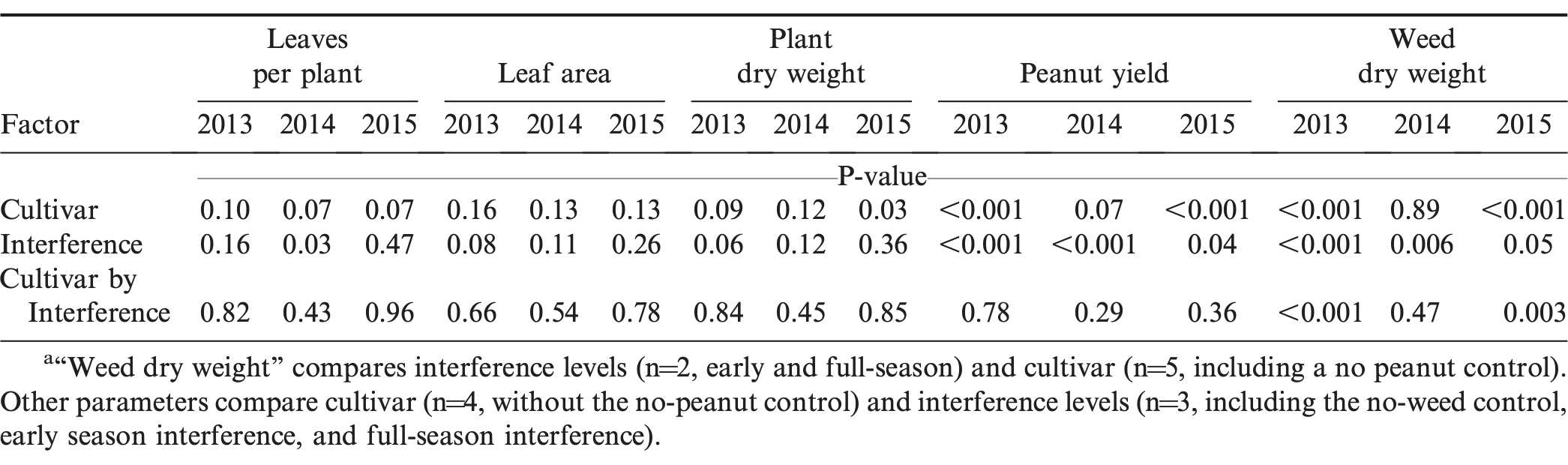
Statistical significance of experimental factors based on analysis of variance for peanut growth and yield parameters and weed dry weight for three years.a
Cultivar, interference, and their interactions were generally not significant for plant dry weight, leaf area, and leaves per plant during all three years (Table 1). A yield response to cultivar was observed in 2013 and 2015, with yields in the order of Georgia-06G ≥ TUFRunner 727 > Bailey ≥ UFT312 (Table 2). TUFRunner 727 performed as well as Georgia-06G in two of the three years. The results indicated that despite their differences in growth habit and canopy architecture, the four evaluated cultivars responded similarly to weed interference (Table 1). Place et al. (2012) compared biomass production of eight Virginia market type peanut genotypes growing under weed-free and weedy conditions, but all genotypes responded similarly to weed interference. They concluded that, at least for Virginia market type peanut, efforts to select cultivars with higher weed suppression and competition tolerance might have little value as part of an integrated weed management approach due to the limited genetic variability for these traits.
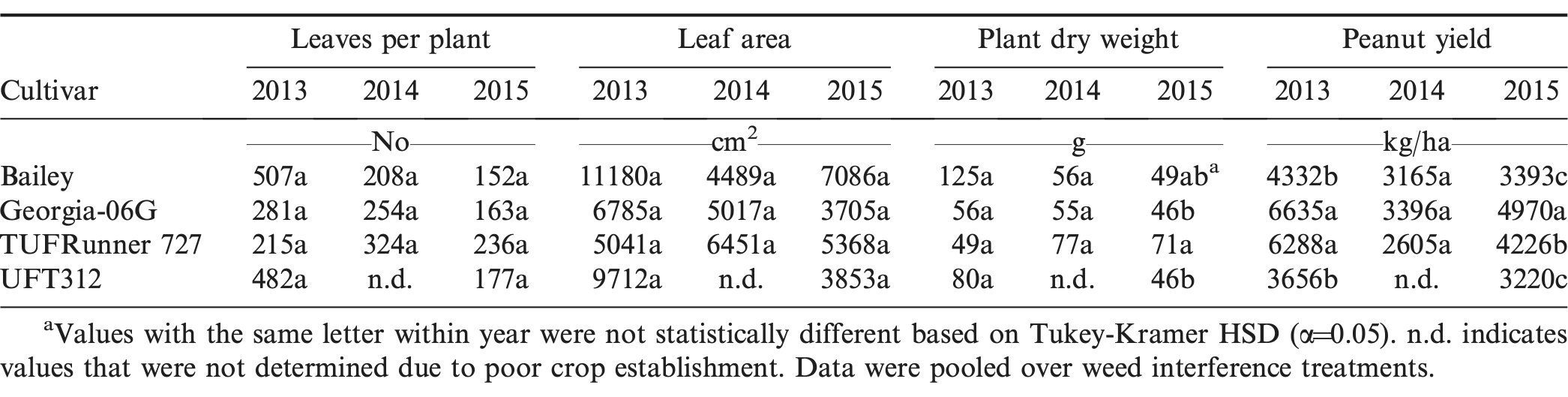
Peanut growth parameters, yield, and weed dry weight for four cultivars grown during 2013 to 2015 in Jay, FL.
Weed interference level significantly affected peanut yield during all three years (Table 1). Weed biomass production was affected by the interaction between cultivar and interference in 2013 and 2015 (Table 1). However, this interaction was mainly due to the magnitude of the differences between the no-peanut weedy control under full-season interference and the cultivar treatment in the same and other interference conditions. When cultivars were compared within interference conditions, they suppressed weed growth similarly (data not shown). For this reason, only main factors are discussed. As weed interference duration increased, weed biomass increased (Figure 2) and peanut yield decreased (Figure 3), confirming that weed populations were high enough to negatively impact peanut growth, development, and yield. In 2013 and 2015, all cultivars suppressed weed growth more than 76% (data not shown). However, in 2014, there was no weed suppression when compared to the peanut free control (P>0.93). Weather conditions (temperature, solar radiation, and total rainfall) were similar during the three years of the study (Figure 1), but fall armyworm ( Spodoptera frugiperda [J.E. Smith]) infestations during 2014 exceeded threshold and required insecticide applications in all plots. The heavy infestation is thought to have reduced weed biomass that year especially in the no peanut treatments (Figure 2). Although weed growth was considerably higher in 2015 than in 2014 (Figure 2), peanut yield reductions due to interference level were lower in 2015 than in 2014 (P<0.0001; Figure 3). These particular results illustrate the complexity of interference between peanut and weeds.
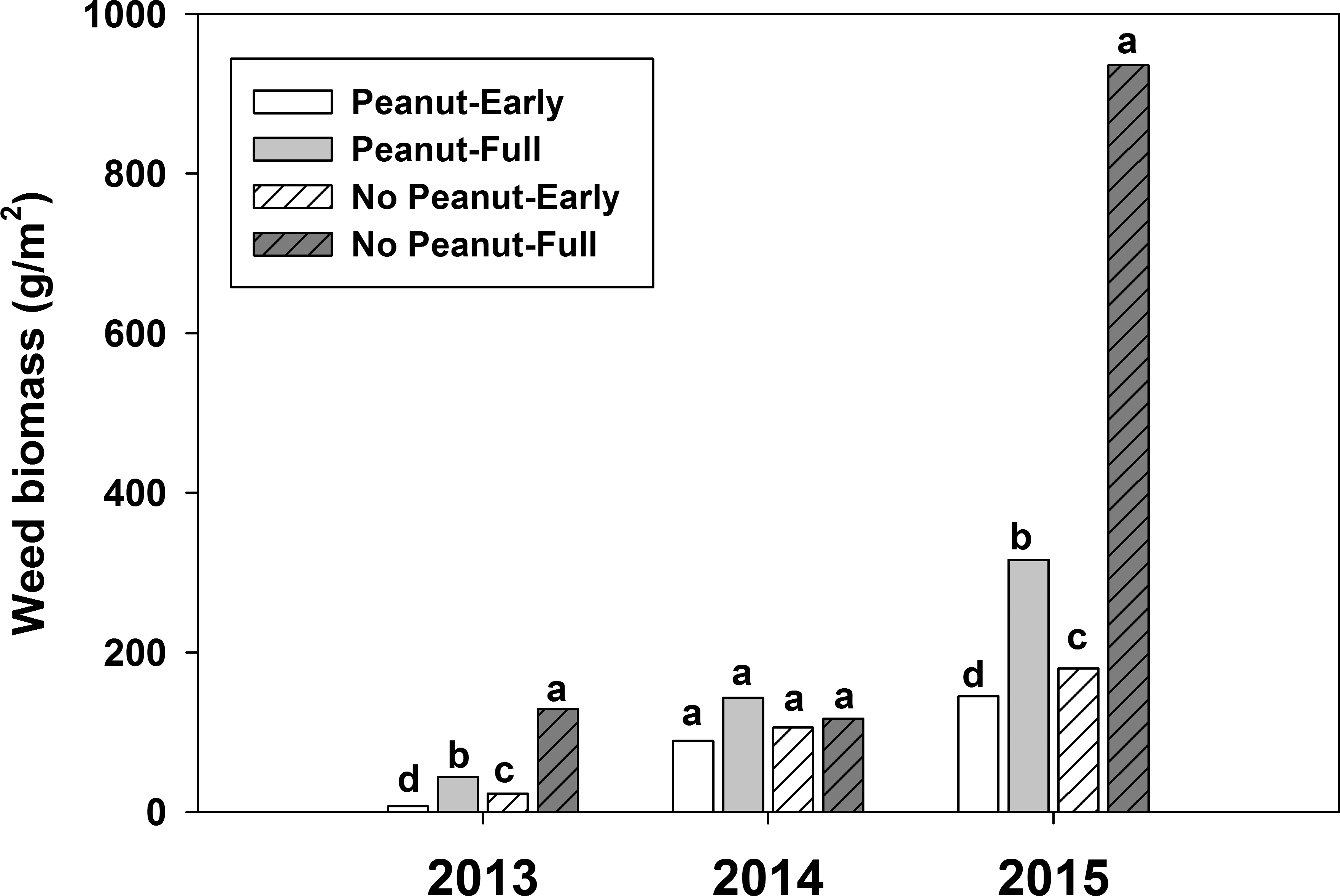
Weed biomass production with and without early and full-season interference with peanut during 2013 to 2015 in Jay, FL. Columns with the same letter within year were not statistically different based on Tukey-Kramer HSD (α=0.05).
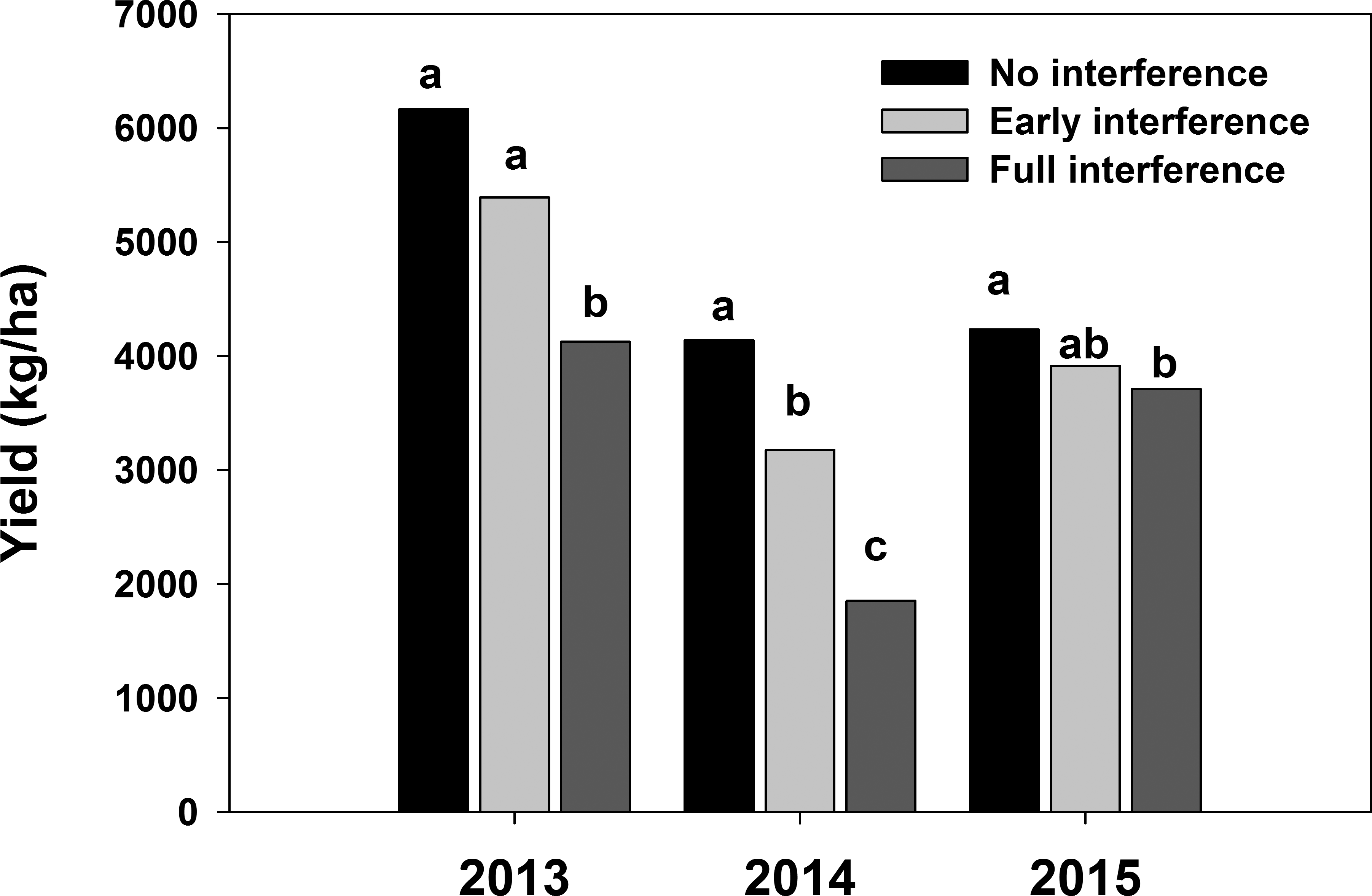
Peanut yield in response to no, early and full-season weed interference during 2013 to 2015 in Jay, FL. Columns with the same letter within year were not statistically different based on Tukey-Kramer HSD (α=0.05).
Weed interference increased peanut main stem height in the order of full-season interference > early season > no weed interference in 2013 and 2014 (Table 3). Canopy height ranking between cultivars was not affected by interference, and Bailey was the tallest cultivar and UFT312 the shortest regardless of interference duration. It is worth noting that in 2015, there was no interaction between peanut cultivar and weed interference for canopy height and width (Table 3). This is an indirect effect of all cultivars being considerably taller in 2015 compared to the other two years (Table 3). As shown in Figure 2, weed populations were highest in 2015, which possibly forced all cultivars to maximize stem elongation to compete against weeds (Ballaré and Casals, 2000; James et al., 1988).
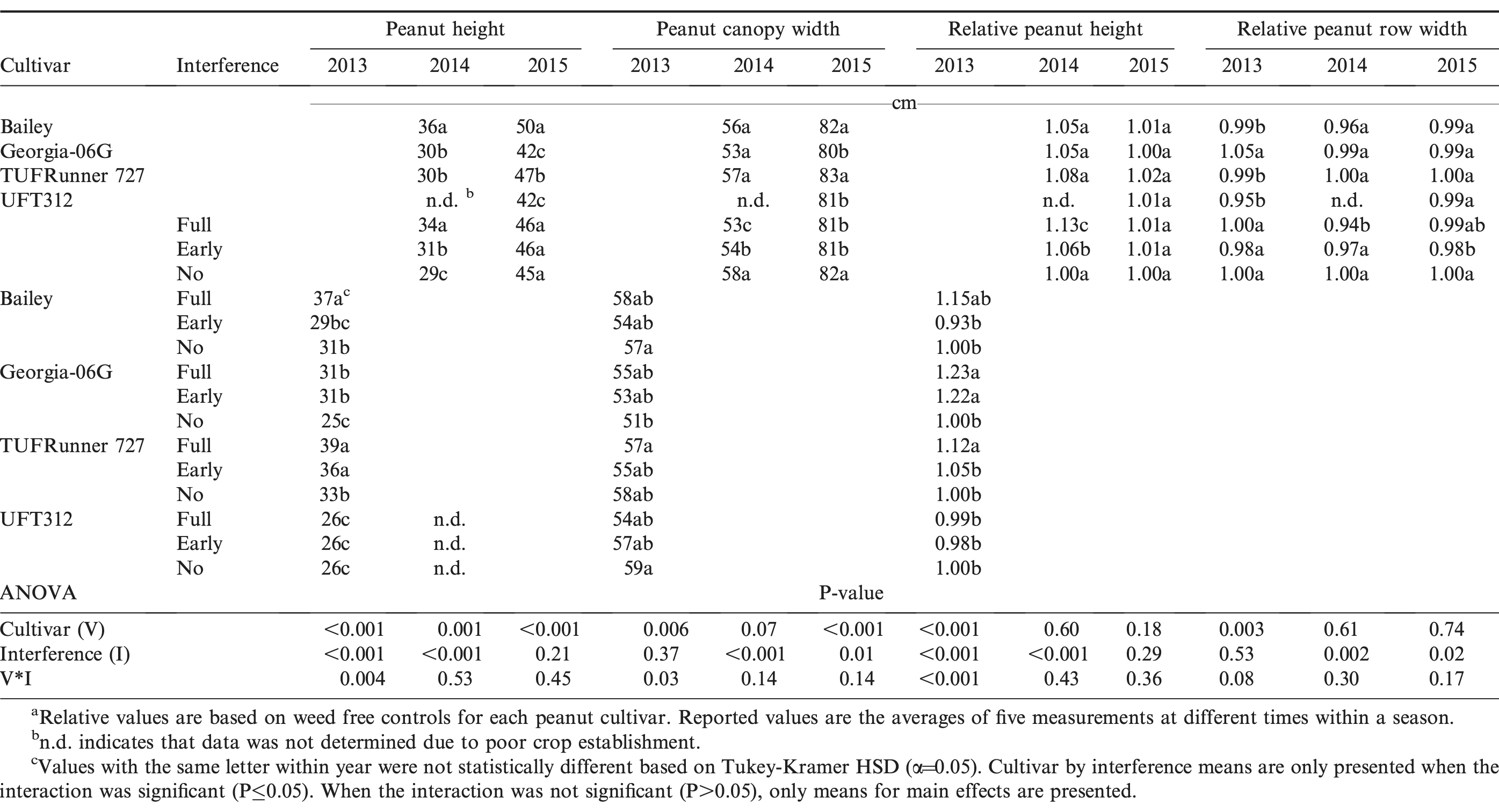
Average absolute and relative peanut height and canopy width based on a repeated measurement analysis for four cultivars and three weed interference durations during 2013 to 2015 in Jay, FL.a
No clear differences were observed between cultivars for relative change in height and width in response to interference (Table 3). A cultivar by interference interaction for peanut height and canopy width was only significant in 2013 (Table 3). In that year, Georgia-06G had the greatest peanut height response to weed interference, while UFT312 did not respond. However, in 2014 and 2015, the lack of a cultivar by interference level interaction indicates that cultivars responded similarly to weed interference, regardless of the differing growth habits of the cultivars chosen for this study.
Peanut yield was inversely related to weed interference duration during the three years of the study (Figure 3). However, the dry weight of peanut plants was not affected by weed interference during all three years (Figure 4). These results indicated that commercial peanut cultivars favor vegetative over reproductive growth while experiencing weed interference even when that interference affects peanut growth.
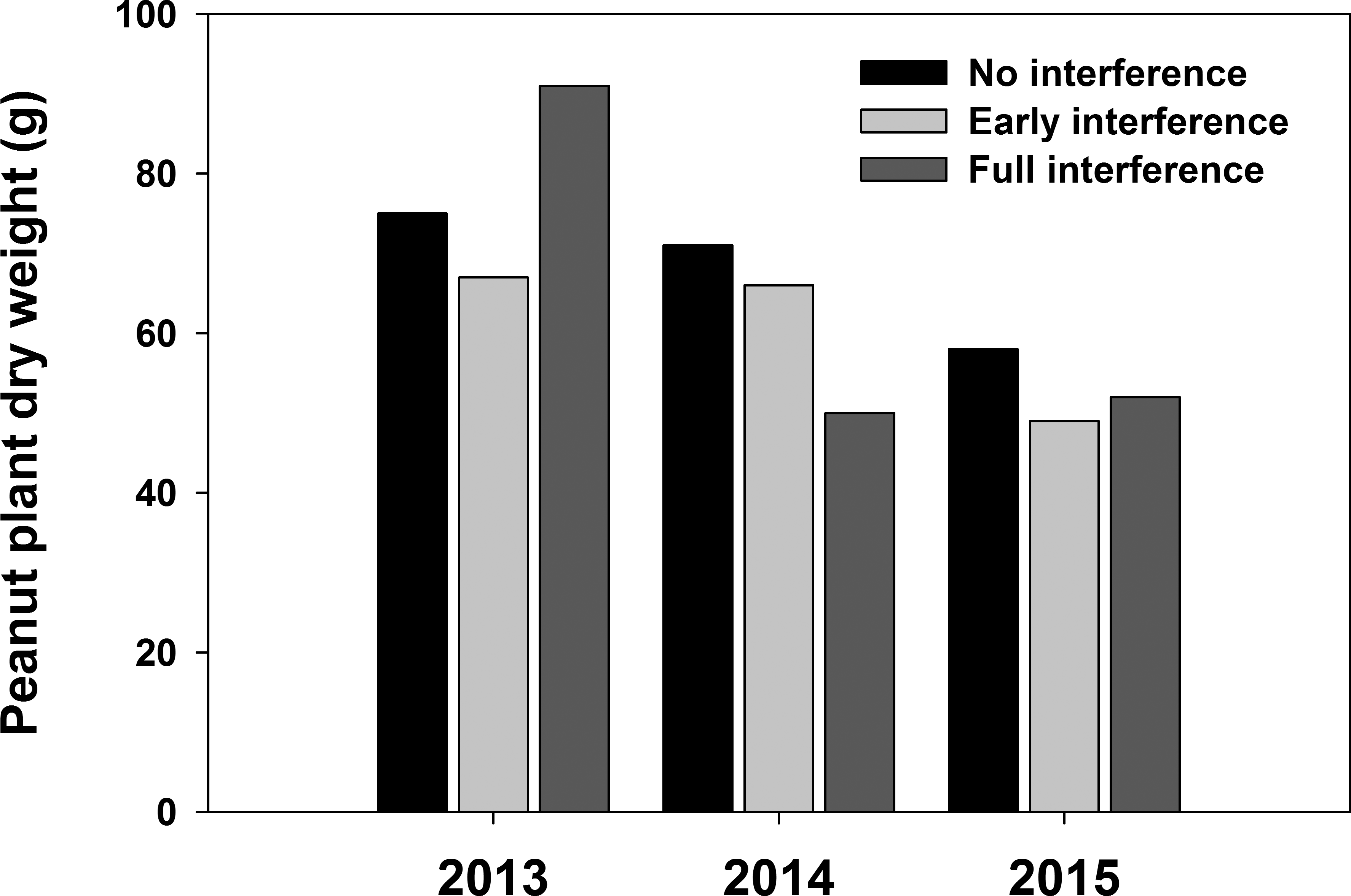
Peanut plant dry weight in response to no, early and full-season weed interference during 2013 to 2015 in Jay, FL. There were no differences between interference treatments based on Tukey-Kramer HSD (α=0.05).
Agostinho et al. (2006) reported that peanut genotypes differed in ability to compete with weeds, and the most evident response was not only yield loss but also a reduction in kernel size when compared with weed-free conditions. Our findings suggest that peanut weed interference tolerance could be improved by identifying lines that would protect yield by favoring photoassimilate translocation to the kernel rather than to above ground tissue during weed interference. Although this strategy might reduce leaf area, this may not necessarily reduce peanut weed suppressive ability (Bussan et al., 1997), especially if the elongation response to weed interference is maintained or increased. Hoad et al. (2008) proposed that crop competitive ability depends not only from the intrinsic weed suppressive ability of the crop based on its morphological characteristics, but also on the sensitivity of the crop to weed interference, and how this sensitivity modulates changes in crop growth to suppress weeds and maintain yield. Therefore, efforts to increase competitive tolerance to weeds should emphasize experimental approaches that allow characterization of peanut sensitivity to weed interference. However, it might be necessary to evaluate peanut types other than those studied in the present research and by Place et al. (2012) to increase the likelihood of finding genetic material with higher potential for weed suppression and interference tolerance
Acknowledgements
We thank Sharon Howell, Michael Dozier, Rocio van der Laat, and Mark Gomillion for technical support. This project was supported by Florida Peanut Producers and the United States of America Department of Agriculture Project 1000851 FL-JAY-005272.
Literature Cited
F.H., Agostinho, R Gravena, P.L.C.A Alves, T.P Salgado, and E.D Mattos (2006). The effect of cultivar on critical periods of weed control in peanuts. Peanut Sci 33 : 29 - 35 .
I.K.S., Andrew, J Storkey, and D.L Sparkes (2015). A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Res 55 : 239 – 248 .
C.L., Ballaré, and J.J Casals (2000). Light signals perceived by crop and weed plants. Field Crop. Res 67 : 149 – 160 .
J.C., Barbour, and D.C Bridges (1995). A model of competition for light between peanut ( Arachis hypogea ) and broadleaf weeds. Weed Sci 43 : 247 – 257 .
W.D Branch, (2007). Registration of ‘Georgia-06G’ peanut. J. Plant Reg 1 : 120 .
A.J., Bussan, O.C Burnside, J.H Orf, and K.J Puettmann (1997). Field evaluation of soybean ( Glycine max ) genotypes for weed competitiveness. Weed Sci 45 : 31 – 37 .
Gorbert, D.W and B.L Tillman 2013 TUFRunner Brand ‘727’ peanut . U.S. P.V.P. No. 201300199.
E.W., Hauser, and G.A Buchanan (1981). Influence of row spacing, seeding rates, and herbicide systems on the competitiveness and yield of peanuts. Peanut Sci 8 : 74 – 81 .
E.W., Hauser, G.A Buchanan, and W.J Ethredge (1975). Competition of Florida beggarweed and sicklepod with peanuts: I. Effects of periods of weed-free maintenance or weed competition. Weed Sci 23 : 368 – 372 .
Heap, I 2014 Herbicide resistant weeds Integrated Pest Management . Springer. p 281 – 301 .
S., Hoad, C Topp, and K Davies (2008). Selection of cereals for weed suppression in organic agriculture: a method based on cultivar sensitivity to weed growth. Euphytica 163 : 355 – 366 .
T.G., Isleib, S.R Milla-Lewis, H.E Pattee, S.C Copeland, M.C Zuleta, B.B Shew, J.E Hollowell, T.H Sanders, L.O Dean, K.W Hendrix, M Balota, and J.W Chapin (2011). Registration of ‘Bailey’ peanut. J. Plant Reg 5 : 27 – 39 .
K.L., James, P.A Banks, and K.J Karnok (1988). Interference of soybean, Glycine max , cultivars with sicklepod, Cassia obtusifolia . Weed Technol 2 : 404 – 409 .
J.L., Jannink, J.H Orf, N.R Jordan, and R.G Shaw (2000). Index selection for weed suppressive ability in soybean. Crop Sci 40 : 1087 – 1094 .
A., Légère, and M.M Schreiber (1989). Competition and canopy architecture as affected by soybean ( Glycine max ) row width and density of redroot pigweed ( Amaranthus retroflexus ). Weed Sci 37 : 84 – 92 .
G.K McDonald, (2003). Competitiveness against grass weeds in field pea genotypes. Weed Res 43 : 48 – 58 .
G.T., Place, S.C Reberg-Horton, and D.L Jordan (2010). Interaction of cultivar, planting pattern, and weed management tactics in peanut. Weed Sci 58 : 442 – 448 .
G.T., Place, S.C Reberg-Horton, D.L Jordan, T.G Isleib, and G.G Wilkerson (2012). Influence of Virginia market type genotype on peanut response to weed interference. Peanut Sci 39 : 22 – 29 .
P.R., Watson, D.A Derksen, and R.C van Acker (2006). The ability of 29 barley cultivars to compete and withstand competition. Weed Sci 54 : 783 – 792 .
E.J., Williams, and J.S Drexler (1981). A non-destructive method for determining peanut pod maturity. Peanut Sci 8 : 134 – 141 .
Notes
- Asst. Prof. and Asst. Prof., West Florida Research and Education Center and Agronomy Department, University of Florida, Jay, FL 32565 [^]
- Assoc. Prof., North Florida Research and Education Center and Agronomy Department, University of Florida, Marianna, FL 32446 [^] *Corresponding author (rglg@ufl.edu)
Author Affiliations