

Introduction
Salmonella is a leading agent of foodborne illness, accounting for thousands of foodborne illness outbreaks reported each year. The Centers for Disease Control and Prevention (CDC) estimates that Salmonella causes approximately 1.0 million illnesses per year in the United States (Scallan et al., 2011), amounting to a conservative annual average economic burden of approximately $4.4 billion (Scharff, 2012). Water activity (aw), where aw = P/P0, and P = partial pressure of water over the sample at equilibrium and P0 = equilibrium pressure of pure water at the same temperature, is a critical concept in microbial safety, as the minimum aw for growth of most bacteria is approximately 0.87. Foods with an aw below 0.85 are commonly classified as low-water activity, and these include roasted peanuts (Beuchat et al., 2013). Although Salmonella contamination has traditionally been associated with high-water activity foods such as eggs and poultry, several recent outbreaks have involved lower-aw foods such as almonds, peanut butter, powdered milk, cereal, chocolate, dry pet food, and various nuts (Beuchat et al., 2013). Salmonella growth generally occurs at aw between 0.94 and 0.99 (Bell and Kyriakides, 2009); however, it can survive at lower aw, and may be difficult to inactivate (Farakos et al., 2013). Survival of Salmonella is a concern in low-aw foods since infection by Salmonella may occur with ingestion of even a single cell, depending on the health of the consumer (Food and Drug Administration, 2012).
Until recent years, nuts have been considered to be of low risk for Salmonella contamination due to low aw. Although Salmonella does not generally grow on nuts, it can survive on nut surfaces for periods as long as a year, and develops increased resistance to thermal treatment over time (Danyluk et al., 2007). One of the largest food recalls in U.S. history was due to an outbreak linked to Salmonella in peanut products causing 714 documented illnesses and 9 deaths, leading to 3,900 products being recalled by 200 companies (Wittenberger and Dohlman, 2010). In addition to peanut butter, peanuts are used as an ingredient in a wide variety of products, including, snack foods and confections (Chang et al., 2013). Many small companies buy peanuts from larger manufacturers who produce bulk quantities of peanuts to distribute to numerous clients (Wittenberger and Dohlman, 2010).
Salmonella contamination can occur at any point in the production of foods and is generally traced back to human or animal sources. Eliminating the risk of Salmonella contamination at the manufacturing level involves proper control and hygiene during all stages of processing, including raw materials handling, roasting, post-roast processing and packaging (Bell and Kyriakides, 2009). Prevalence of Salmonella on raw, grade samples of peanuts from all production areas in 2008 to 2010 was estimated at 2.33% and the concentration levels of Salmonella in positive samples, as determined by a most-probable-number assay (MPN), were <0.03 to 2.4 MPN/g (Calhoun et al., 2013). In a separate study, Salmonella was found in 0.67% (68/10,162 samples) from raw, runner peanuts from 2009, 2010 and 2011sampled at labs located in both GA and TX (Miksch et al. 2013). No difference in prevalence data was observed for the two growing regions and concentration levels of Salmonella in positive samples averaged 1.05 MPN per 350 g.
The FDA recommends that peanut processors use a process that is validated to reduce Salmonella on peanuts, e.g., 5 logarithm of colony forming units/g (5-log CFU/g) (Food and Drug Administration, 2009). Commercial testing laboratories that perform roaster validations currently use a Salmonella surrogate, Enterococcus faecium (personal communication, Deibel Laboratories, Inc.). This organism was used in almond processing validations (Almond Board of California 2007a, 2007b) and in recent studies on peanuts was found to be slightly more heat tolerant than Salmonella (Sanders and Calhoun, 2014). Roasting is used as the critical control point in most peanut processing operations. The two most common types of roasting are oil roasting and dry roasting. Studies have been performed to investigate Salmonella reduction in the thermal processing of various nuts, and in the later processing operations for several peanut-based products, such as peanut butter (Shachar and Yaron, 2006; Ma et al., 2009 Sanders and Calhoun, 2014). Research on oil roasting of peanuts (Sanders and Calhoun, 2014) and almonds (Du et al, 2010) suggested that 5-log CFU/g or greater reductions were expected at normal oil roasting temperatures (150–160 C) because of the higher thermal conductivity of oil compared to air. Dry roasting is the most widely used roasting process in the peanut industry. Some information is available regarding reduction of Salmonella during dry roasting of almonds (Bari et al., 2009), sesame (Torlak et al, 2013), pecans (Beuchat and Mann, 2011), and peanuts (Sanders and Calhoun, 2014). Sanders and Calhoun (2014) reported an examination of E. faecium and a cocktail of 7 different Salmonella cultures inoculated onto peanuts which were then subjected to a series of both dry and oil roasting temperatures for different times. They utilized 25 g samples and bed depth and air flow were not considerations in the studies. The data reported demonstrated that at every time and temperature, E. faecium was slightly more heat tolerant than Salmonella. These data demonstrated that E. faecium was an excellent surrogate for Salmonella as it was previous shown to be for almonds.
Given the lack of information available on reduction of Salmonella on peanuts in industrial dry-roasting processes which include temperature, time, bed depth, air flow, and air flow direction, the objectives of this study were; (i) to estimate the effectiveness of various industry-typical dry roasting conditions in reduction of Salmonella; (ii) to determine if Salmonella reduction depends on location in the bed of peanuts in the dry roaster; (iii) to determine if Salmonella reduction correlates to any physical roasted peanut properties.
Materials and Methods
Peanuts
A single lot of medium runner-type peanuts (Georgia O6G cultivar) obtained from a commercial processor was used for all studies. Georgia O6G (Branch, 2007) was selected as this is the most common cultivar of peanut grown in the Southeastern United States, accounting for approximately 77 percent of seed acres in 2012 (Brandon, 2013). All tests were conducted using whole, unblanched peanuts. Initial moisture of the peanuts was 6.1% on a wet weight basis (wwb).
Inoculation and Quantification
Inoculation of the peanuts was performed by Deibel Laboratories (3530 NW 97th Blvd, Gainesville, FL 32606) using a surrogate cocktail combination of E. faecium ATCC 8459 and E. faecium ATCC 35667 as utilized in studies conducted by the Almond Board of California (2007a, 2007b) and Sanders and Calhoun (2014). All inoculum cultures were grown in tryptic soy broth (BD Diagnostics, 7 Loveton Circle, Sparks, MD 21152). The nuts were inoculated and stabilized back to their original moisture levels using the protocol established by the Almond Board of California (2007b).
The inoculation procedure involved diluting unroasted peanut samples at a 1∶10 ratio with sterile Butterfield's Phosphate Buffer (BD Diagnostics, 7 Loveton Circle, Sparks, MD 21152) and mixing for 2 minutes employing a Stomacher Lab Blender (Fisher Scientific, 100 Technology Dr, Pittsburgh, PA 15219). The samples were then held for 3 to 5 min prior to further dilution. The samples were then vigorously shaken by hand a minimum of five times followed by serial dilution in 9 ml tubes of sterile peptone diluent. For each dilution, 0.1 ml aliquots of diluted sample were spread plated onto KF Streptococcus Agar plates (BD Diagnostics, 7 Loveton Circle, Sparks, MD 21152) tempered at ambient temperature for at least 4 hours. Plates were incubated in a Fisher Scientific Isotemp Lab Incubator (81 Wyman St., Waltham, MA 02454) at 35 C for 48 hours.
Inoculated samples were enumerated prior to shipment to confirm that the inoculation level was at least 6.5-log CFU/g. After samples for determination of inoculation levels had been removed, two 25 gram samples of the inoculated peanuts were scattered on aluminum mesh racks with the nuts separated and exposed to a heat treatment of 138 C for 15 minutes in a Fisher Scientific Isotemp 851F oven (81 Wyman St., Waltham, MA 02454). After heat treatment, the samples were plated as above. A reduction of less than 2.5-log CFU/g after the prescribed heating trial indicated the appropriate heat resistance of the surrogate cocktail (Almond Board of California 2007b).
Inoculated peanuts were held at 4 C after overnight shipment to the roasting location. Over the course of these studies, un-roasted, inoculated samples were tested to ensure that the initial level of inoculation was consistent for all trials and varied from 6.5 to 7.7-log CFU/g. Inoculation levels were verified prior to a given roast treatment and resulting log reduction levels were calculated accordingly.
During the roasting trials, 25 g samples of inoculated peanuts were equilibrated to ambient temperature in a cotton mesh steeping bag (commonly used in home beer brewing), which was positioned in the desired location (top, middle or bottom) of the peanut bed. Sterile conditions were used immediately after roasting and cooling protocols, to refrigerate samples that were shipped overnight in insulated coolers with icepacks for enumeration of surviving E. faecium.
Treated samples were received at the Deibel Laboratory intact and under refrigeration. All treated samples and controls were enumerated twice on KF Streptococcus according to the protocol outlined above. After 48 hours at 35 C, plates were enumerated and the log CFU/g values of these counts were calculated. From these log values, minimum log reductions were calculated from the two enumeration results.
Roasting
Roasting was conducted in a laboratory scale roaster simulator (Figure 1) in which temperature, time, airflow speed, airflow direction and bed depth were regulated. The batch-type roaster consisted of a centrifugal fan for airflow, an electric heater and a rectangular tray which held 2 to 3 kg of peanuts depending on selected conditions. The square base of the tray (0.044 m2) was uniformly perforated. Temperatures (149, 163, 177, and 191 C), bed depths (38, 75, and 113 mm) and air flows (1.0, and 1.3 m/s) were variously combined to simulate industrial roaster conditions. Air temperature was measured and recorded manually with type J thermocouples from Omega Engineering (One Omega Drive, P.O. Box 4047, Stamford, CT 06907-0047) positioned above and below the tray containing peanuts. Airflow through the tray containing peanuts was measured using an orifice plate in the tray. Dampers in the roaster were used to switch the airflow through the tray between up-flowing air and down-flowing air. Airflow always began as up-flow and was changed to down-flow halfway through the roasting time, which is the most common configuration for continuous dry roasters in the industry. After roasting, the peanuts were cooled to ambient temperature (∼25 C) using ambient up-flowing airflow through the tray of roasted peanuts to simulate the convection cooling commonly incorporated into an industrial dry roaster.
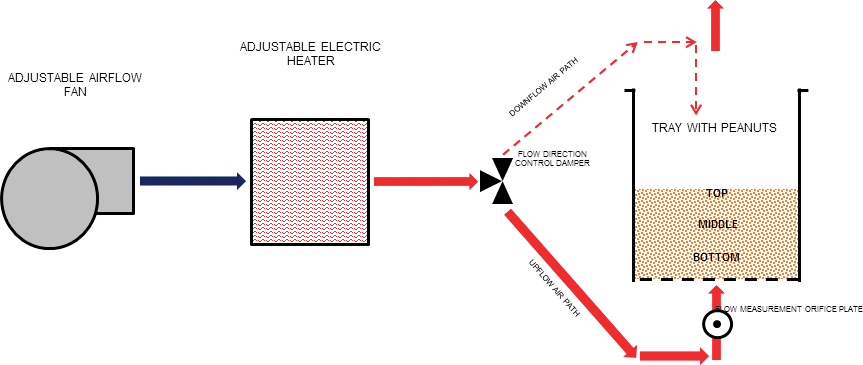
Schematic of laboratory roaster.
Bed and Peanut Temperature
Multiple type K thermocouples and a data logger from Omega Engineering (One Omega Drive, P.O. Box 4047, Stamford, CT 06907-0047) were used to measure and collect temperature data every 5 seconds during each test. Three thermocouples were located in the center of the tray, one at the bottom, one in the middle and one at the top of the bed of peanuts. In addition, a thermocouple was inserted approximately 3 mm into a peanut, which was placed near the middle bed temperature probe. Calibration of the data logger was done by an independent laboratory and checked using boiling water.
Measurement of Moisture, Water Activity and Color
Representative samples of each roast treatment were collected for additional measurements. Peanut moisture content was determined by drying the peanuts in a convection oven at 80 C until no further loss in weight occurred. Moisture content was calculated from wet and dry weights. Water activity was determined using an AquaLab water activity meter from Decagon Devices, Inc. (2365 NE Hopkins Court, Pullman, WA 99163). Roast peanut color was measured using a HunterLab DP-900 Colorimeter from Hunter Associates Lab (11491 Sunset Hills Rd, Reston, VA 20190). Hunter L values (McDaniel et al, 2012) of coarsely ground, blanched (skins removed) peanuts are reported.
Statistical Analysis
All roast treatments (a specified combination of roast temperature, roast time, air flow rate and bed depth) were evaluated in duplicate. Analysis of variance was performed using JMP® Ver. 9.0 (SAS Institute Inc., 100 SAS Campus Drive, Cary, NC 27513). Means were separated using a Student's t test when a significant F-value was obtained.
Results and Discussion
Most continuous, industrial dry roasters (Figure 2) consist of a perforated steel belt that conveys the peanuts through the nut roaster. Peanuts are typically stacked on the conveyor to depths between 50–125 mm (75 mm is a commonly used bed depth). As the conveyor moves the peanuts through the dry roaster, the bed of product is subjected to forced convection airflow at elevated temperatures, typically between 140 and 190 C. Airflow through the bed of product is between 0.7–1.3 m3/s per m2 of bed (expressed as m/s in this study). The dry roaster is divided into independent roaster zones and the direction of the airflow through the bed of peanuts typically alternates between upflow and downflow from one zone to the next. The parameters used in this study (bed depth, process air temperature, airflow, airflow direction) were specifically chosen to simulate the industrial dry roaster conditions described above.
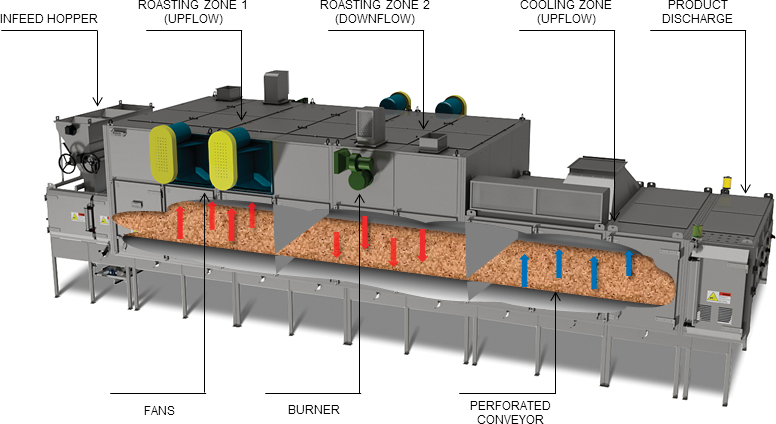
Industrial dry roaster.
Moisture content of the peanuts prior to roasting was 6.1% wwb. After roasting at different time/temperature combinations (constant bed depth of 75 mm and air flow of 1.3 m/s) the highest moisture content observed was 1.7% wwb (Figure 3). At equivalent roast times, moisture content was reduced more at higher roast temperatures. For example, after 12 min roasting, samples prepared at 163 C and 177 C had greater (P<0.05) moisture contents than samples prepared at 191 C, whereas at 16 min roasting, samples prepared at 149 C had greater (P<0.05) moisture content than samples roasted at 163 C. Across these test conditions, linear (R2 ≥ 0.66) trends were observed in moisture loss with increasing roast time at a given roast temperature. Many food processors control the final moisture of their product to ensure that the product is below a target water activity (aw), where aw = P/P0, and P = partial pressure of water over the food sample at equilibrium and P0 = equilibrium pressure of pure water at the same temperature. Water activity is a critical concept in microbial safety, as the minimum aw for growth of most bacteria is approximately 0.87. Foods with an aw below 0.85 are commonly classified as low-water activity, and these include roasted peanuts (Beuchat et al., 2013). Importantly, many low-water activity foods have been implicated over recent years in microbial illness outbreaks, and it is now understood that some bacteria, including Salmonella, can survive for some limited time on low-water activity foods if contaminated (Beuchat et al., 2013). Water activity is also critical in governing various reactions important to final product quality, including lipid oxidation, hydrolytic reactions, non-enzymatic browning and enzyme activity (Fennema, 1996). Water activity for peanuts prior to roasting was approximately 0.73 and after roasting this value ranged from 0.33 to 0.13 (Figure 4). Water activity for roasted peanut samples decreased with increasing roast time at a given roast temperature (Figure 4), although linear trends were not as good as those observed for corresponding moisture content data (Figure 3). This is attributable to water activity data being inherently less accurate/precise for low moisture, high lipid products such as roasted peanuts. Regardless, at equivalent roast times, aw was lower at higher roast temperatures (Figure 4). For all roasted peanut samples, aw was less than 0.35, meaning no roasted peanut would be at risk for microbiological growth if properly packaged with a suitable moisture barrier; however, documenting survival after various roast conditions is critical.
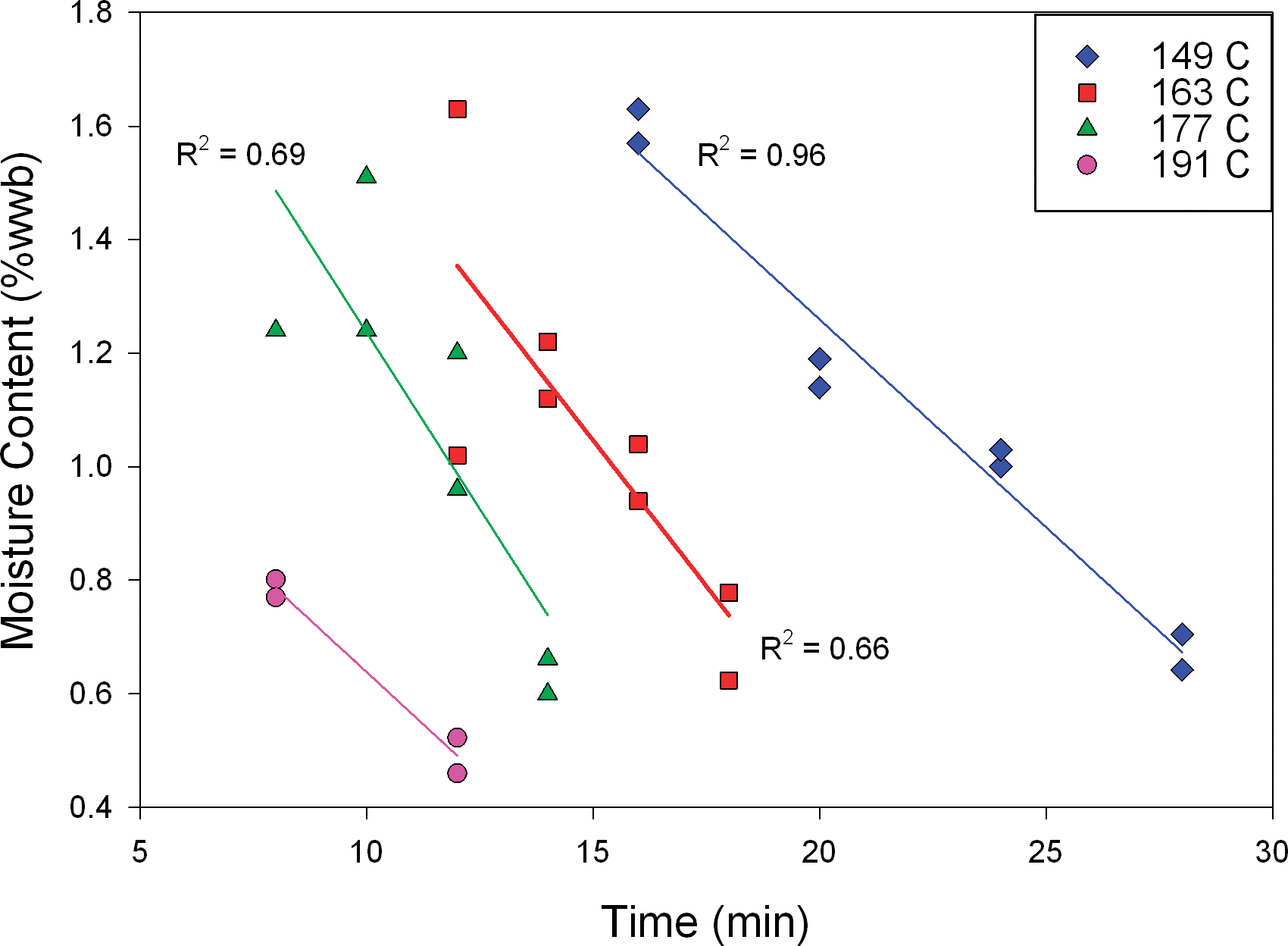
Final moisture content versus roasting time, by temperature, at 75 mm bed depth and 1.3 m/s air velocity.
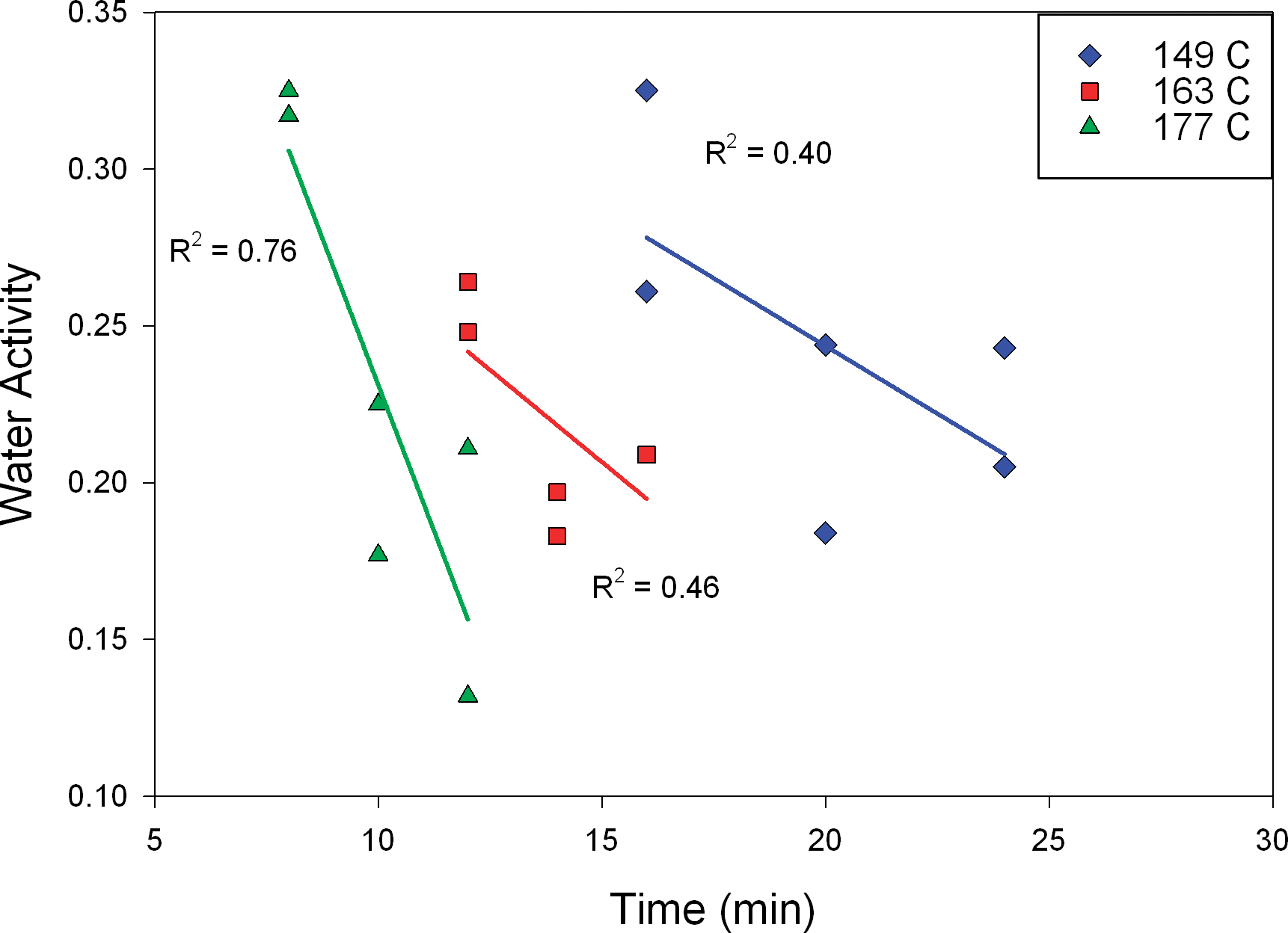
Water activity versus roasting time, by temperature, at 75 mm bed depth and 1.3 m/s air velocity.
During all roasting trials, the air flow direction was up through the peanut bed for the first half of the roasting time and down through the bed for the second half. This protocol simulates industrial roasting procedures in which air flow is alternated as the peanuts progress through the roaster zones. This alternating air flow prevents over roasting of either the bottom, or top, of the bed that would result from unidirectional airflow. Real time temperature data was collected for all roasted samples, and Figure 5 shows a typical temperature plot as measured via thermocouples placed at the top, middle and bottom of the peanut bed. As air flowed up for the first half of each roasting trial, the hot air first contacts the peanuts at the bottom of the bed and the air subsequently loses temperature as it flows upward through the bed, as evident in Figure 5. For this reason, peanuts at the top of the bed are initially subjected to lower temperatures than those at the bottom of the bed. However, half way through the prescribed roasting time, the airflow direction is reversed and the heated air contacts peanuts at the top of the bed first. As evident in Figure 5, the heated air loses energy as it passes down through the product bed and initially cools the peanuts at the bottom of the bed (Figure 5). After roasting, the nuts were cooled using up-flowing air (the industry standard), meaning nuts at the bottom of the bed cooled most rapidly.
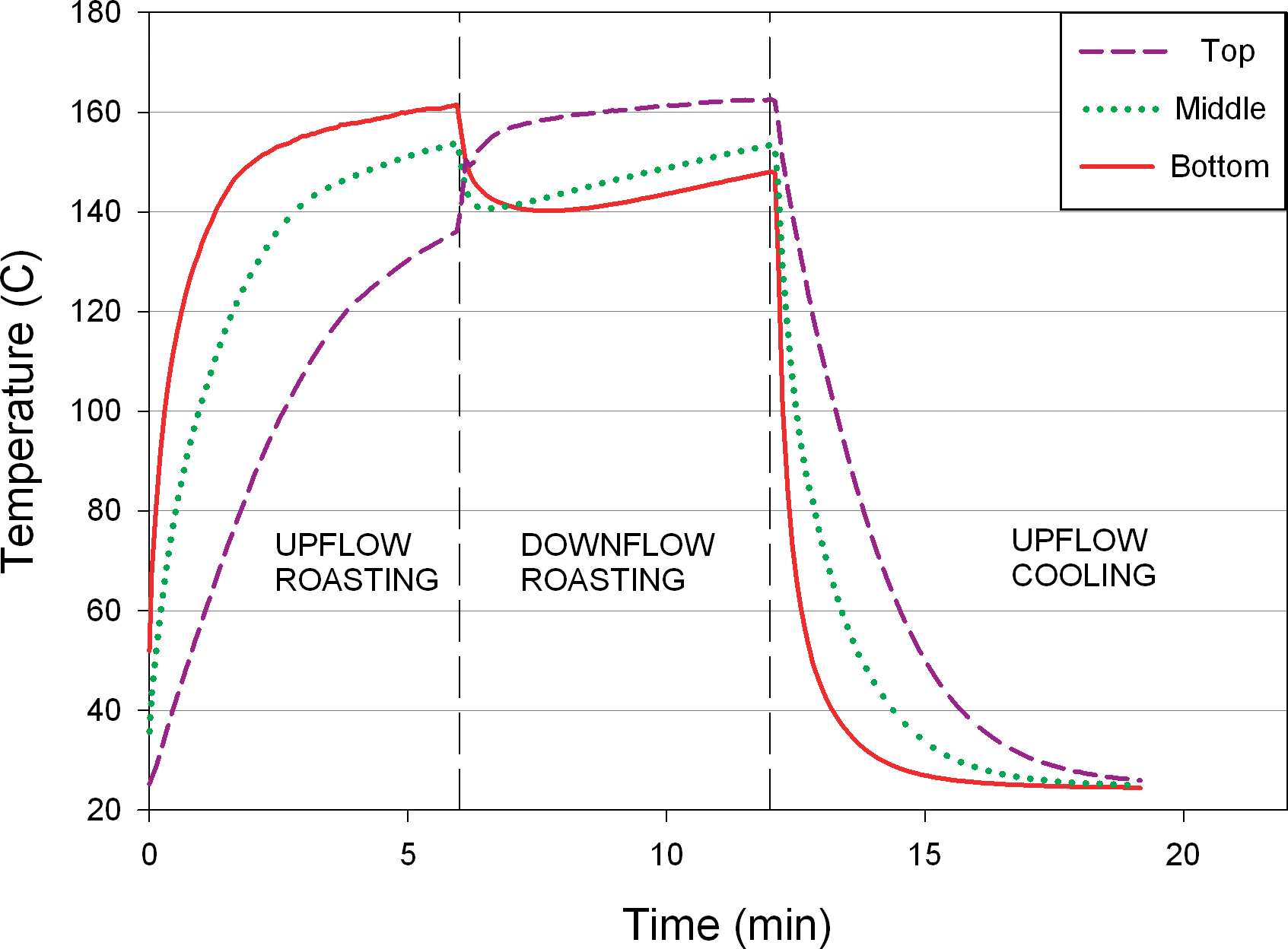
Air temperature at top, middle, and bottom of the bed over time.
The thermocouple measurements in Figure 5 show measurements of the local conditions in the interstices between peanuts and are most representative of the surface temperature of the peanuts. Since Salmonella will likely exist on the surface of peanuts, these measurements most closely represent the conditions to which Salmonella cells are exposed during the roasting process. Based on these temperature measurements, it was evident that thermal inactivation of Salmonella during dry roasting would vary throughout the depth of the product bed. A thermocouple was also embedded inside a peanut for each roast trial to better understand internal peanut temperature during roasting. Figure 6 shows an example of the relationship between the internal temperature of the peanut and the temperature in the interstices around the peanut. The internal temperature lags behind the surface temperature because of the conductive heat transfer through the peanut; however, the internal temperature of the peanut reaches the process air temperature.
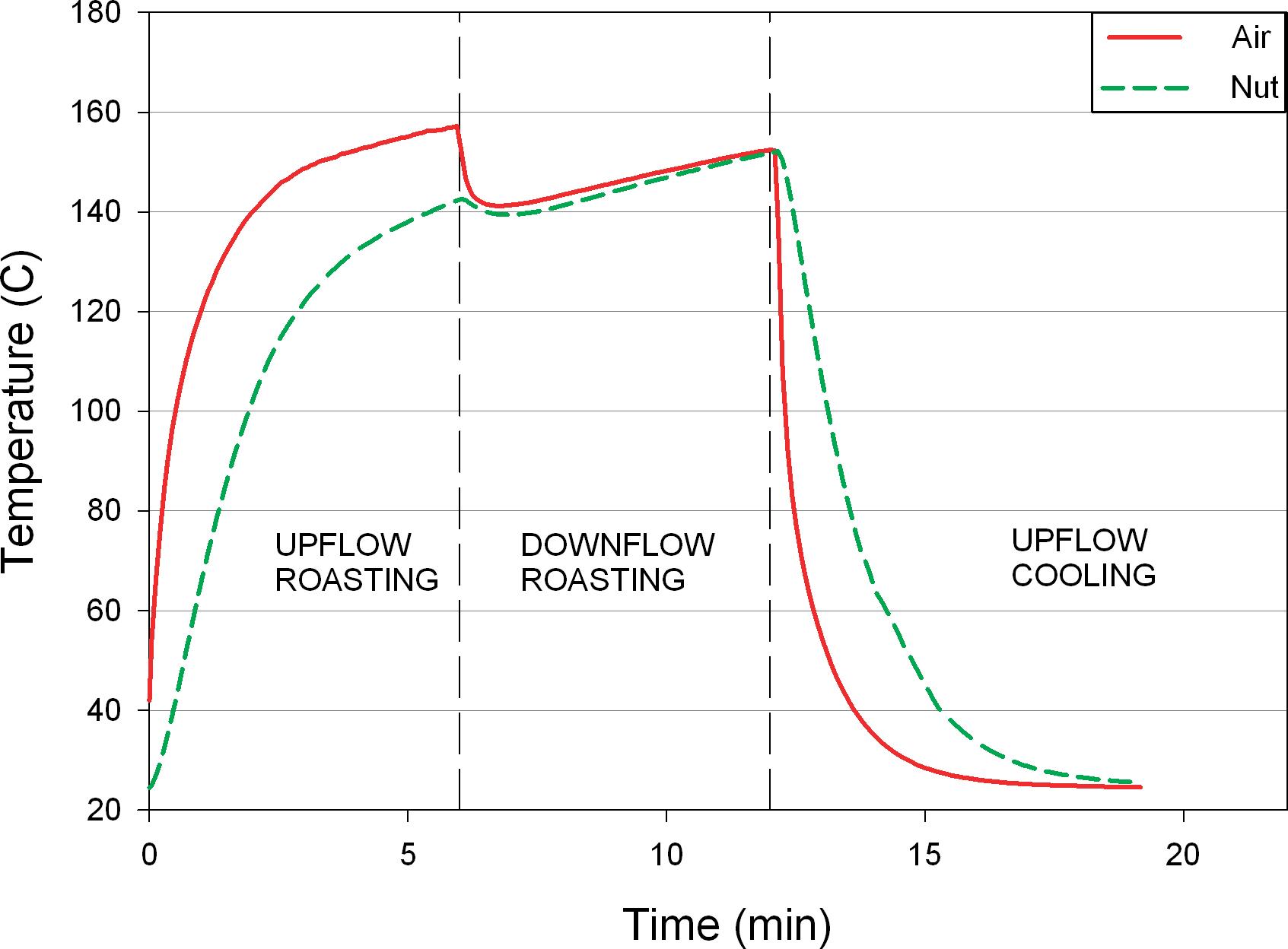
Air temperature compared with internal nut temperature over time.
Salmonella surrogate log reduction was measured at the top, middle and bottom of the peanut bed (75 mm depth) after 14 and 16 min of roasting at 163 C with a constant air velocity of 1.3 m/s (Figure 7). Log reductions were significantly (P<.05) lower at the middle of the bed, compared to the top or bottom, across these conditions (Figure 7). Furthermore, log reduction was greater (P<.05) after 16 min of roasting compared to 14 min, regardless of measurement location (Figure 7). When samples were roasted at 163 C with a bed depth of 75 mm for 16 min, increasing the air flow from 1.0 to 1.3 m/s significantly (P<.05) increased log reduction in the middle and top of the bed, with the greatest increase occurring at the middle of the bed (Figure 8). Data in Figs. 7 and 8 indicated that the middle of the bed always exhibited the lowest (P<.05) level of Salmonella surrogate reduction. The California Almond Board procedures for validation of almond dry roasters recommends “embedding” the inoculated sample in the bed of almonds on the roaster conveyor (Almond Board of California, 2007b), and these results support that approach. Data in Figs. 7 and 8 also show that any change in processing parameters that allow the hot air to more effectively heat the nuts at the middle of the bed will increase Salmonella reduction and decrease variation in Salmonella reduction when comparing reductions at the top, middle and bottom of the bed. Figure 7 shows that longer roasting time increases Salmonella surrogate reductions and ultimately results in more consistent inactivation across the bed depth. Figure 8 shows that higher air flow rates through the bed also increases Salmonella surrogate reduction and results in more consistent inactivation throughout the bed of peanuts. Higher air flow heats peanuts in the middle of the bed more rapidly since the air loses less energy as it travels through the peanut bed. Based on these results, all subsequent Salmonella surrogate inactivation data was collected at the middle of the bed, as this location represents the worst case in terms of inactivation.
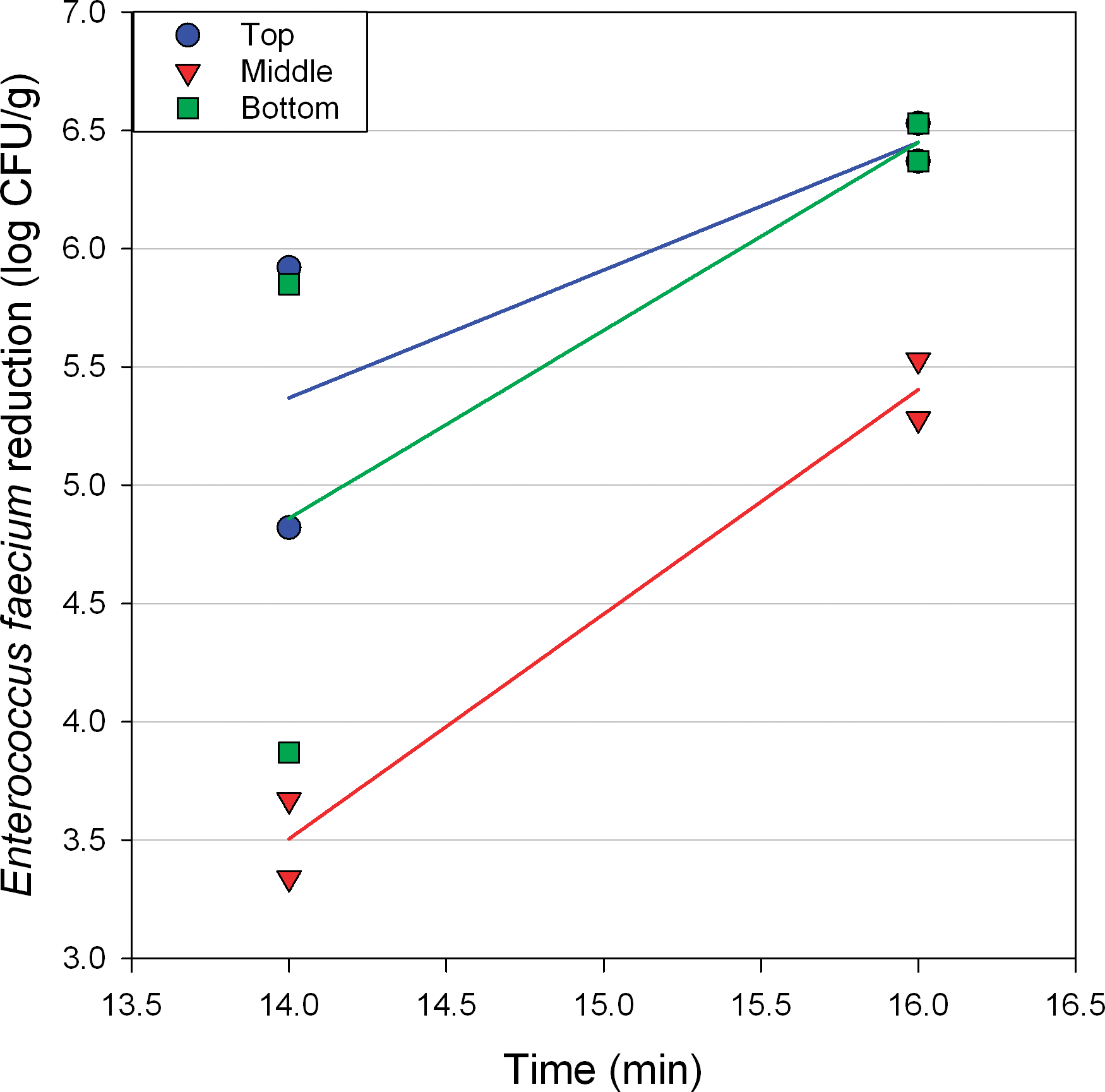
Enterococcus faecium log reduction vs. roasting time at 163 C, 75 mm bed depth, and 1.3 m/s air velocity. Lines are added to better visualize paired data points.
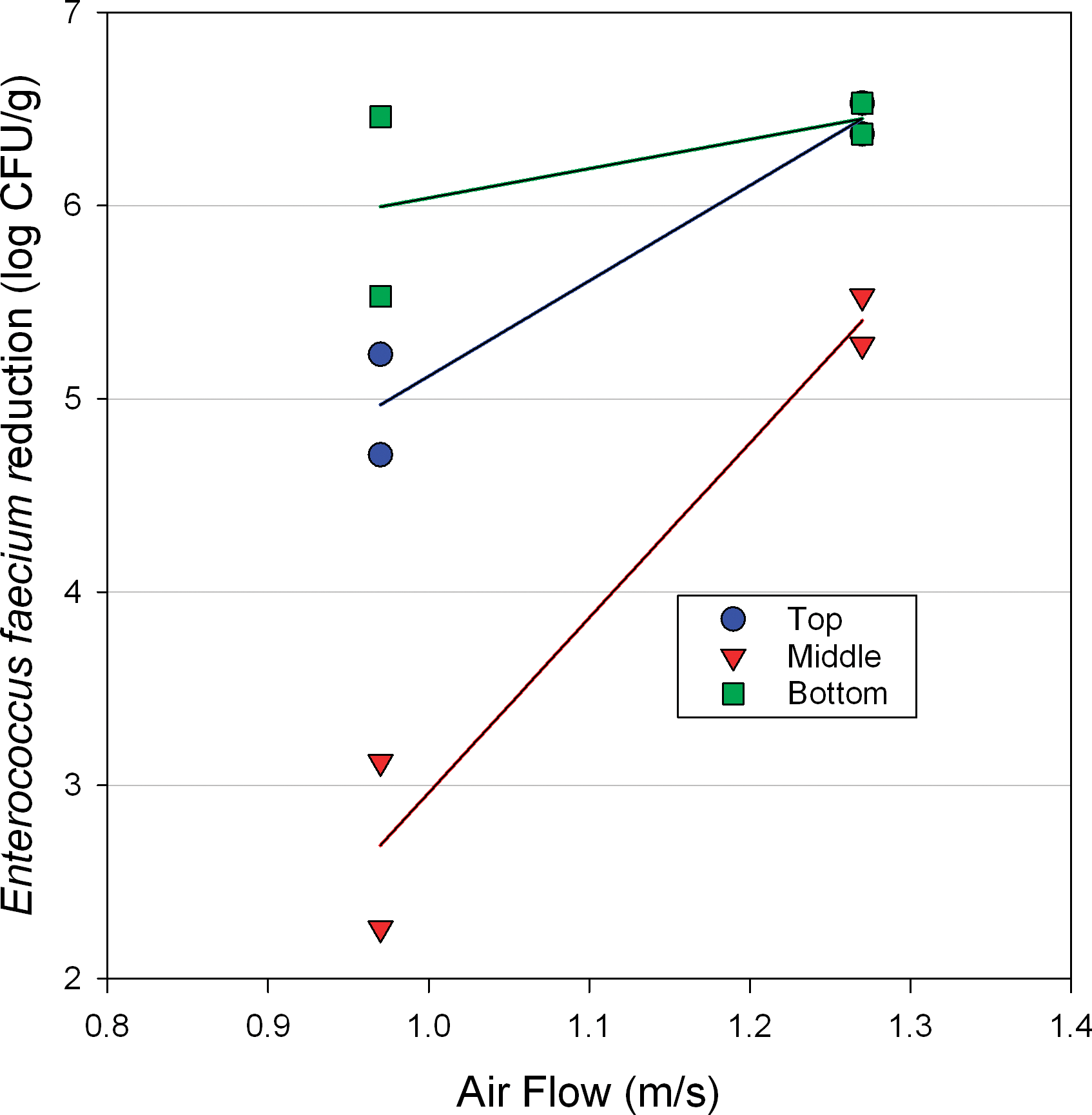
Enterococcus faecium log reduction vs. air velocity at 163 C and 75 mm bed depth with 8 minutes of upflow and 8 minutes of downflow. Lines are added to better visualize paired data points.
Another processing parameter that impacts heating at the middle of the bed is the bed depth. Peanuts were roasted at 163 C for 16 min using 3 bed depths, 38, 75 and 113 mm and log reductions were measured at the middle of the bed (Figure 9). Log reduction was negatively linearly correlated (R2 = 0.67) with increasing bed depth. A deeper bed reduced the capacity of the process air to heat the peanuts in the middle of the bed. Over this limited data set, bed depth impact on log reduction was significant at the P = 0.16 level.
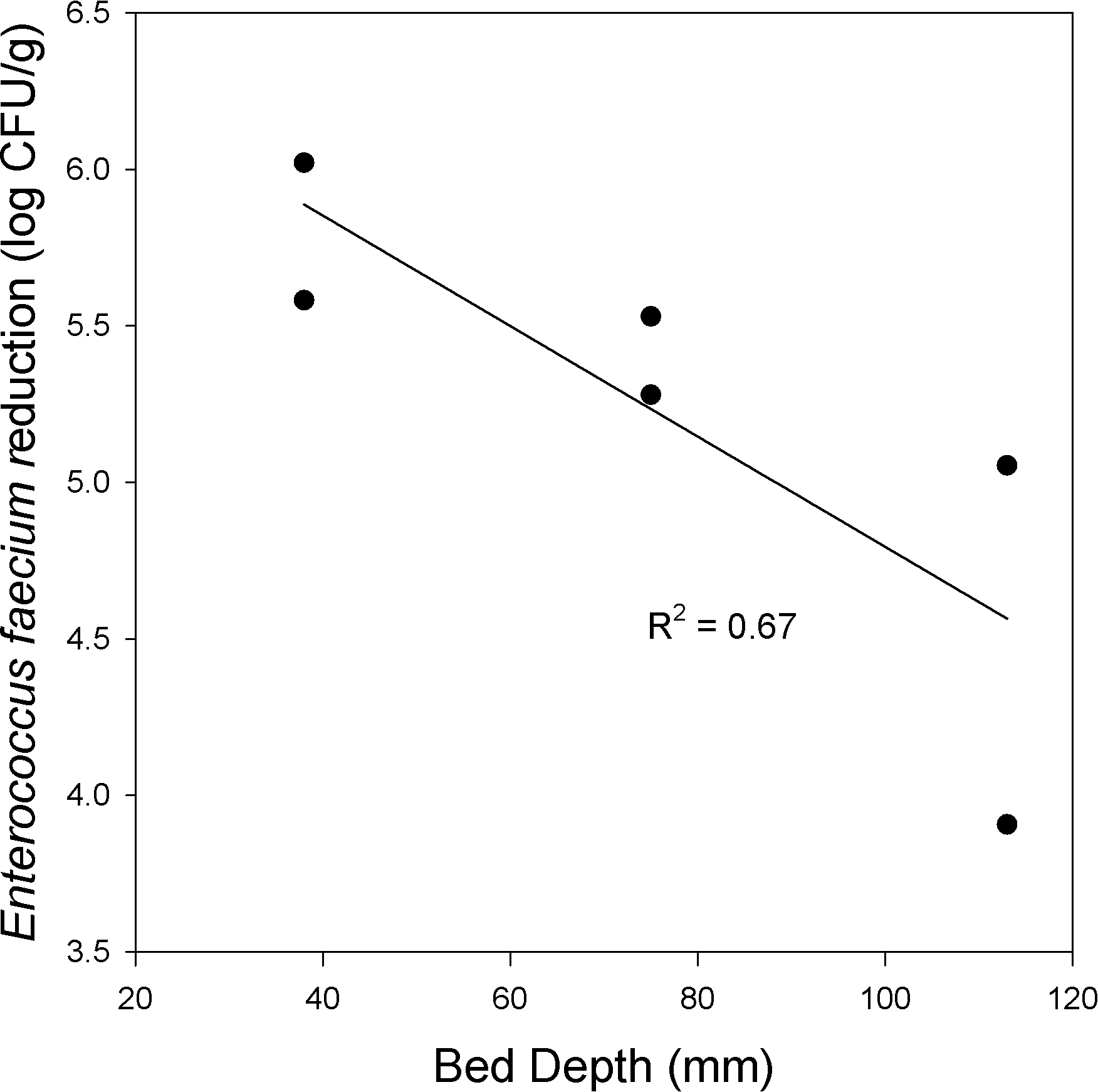
Enterococcus faecium log reduction vs. bed depth at 163 C with 8 minutes of upflow and 8 minutes of downflow.
Figure 10 shows the impact of time and temperature of roasting on Salmonella surrogate log reduction at the middle of the bed (75 mm) with a constant air velocity of 1.3 m/s. Higher temperatures and longer roast times increased inactivation across these conditions, with log reductions generally increasing linearly with roast time at a given roast temperature. The 5-log reduction in CFU/g line is highlighted on this graph since this is a common target in the peanut roasting industry (Food and Drug Administration, 2009). A 5-log CFU/g reduction was achieved during dry roasting at the middle of the product bed under these conditions at 11, 15 and 21 min when roasting at 177, 163 and 149 C, respectively.
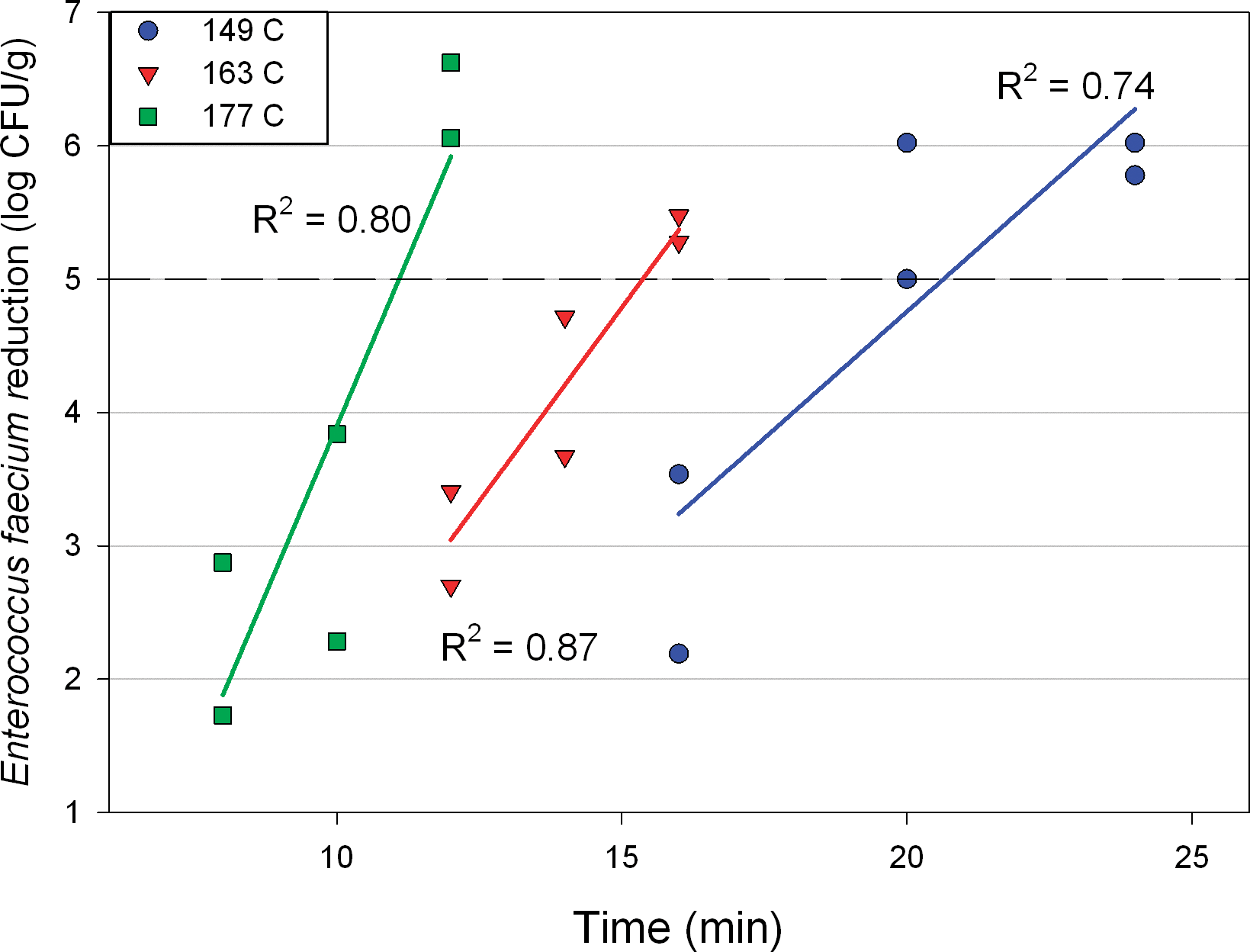
Enterococcus faecium log reduction vs. roasting time, by temperature, at 75 mm bed depth and 1.3 m/s air velocity.
Standard industry practice is to roast peanuts to a target color, as color measurements are rapid, inexpensive, nondestructive and correlate well with final product quality, including flavor, texture, and nutritional composition (McDaniel et al, 2012). Specific L-values from the Hunter Lab scale are commonly targeted in the peanut industry as the L-value corresponds to the relative lightness or darkness of the sample, with 100 and 0 equaling perfectly white and dark, respectively. Figure 11 shows the same log reduction data from Figure 10 plotted versus Hunter-L values of the roasted, ground peanuts. L-values of 53 and 48.5 are industry commonly accepted L values for light and medium colored peanuts, respectively (McDaniel et al, 2012). This graph indicates most peanuts roasted to an L-value of 53 and darker had more than a 5-log CFU/g reduction. Further study is needed to understand and bolster this finding, but this data suggests a good correlation between E. faecium reduction (and thus Salmonella reduction) and Hunter-L color, regardless of roast conditions. As color darkens with both increasing time of roast and increasing temperature of roast, roast color is a good indicator of total heat exposure, which could explain the good correlation observed in Figure 11. However, since roast color is well established to be a function of seed maturity and production location, among other factors, further data is required to understand this correlation and processors should not use roast color as an indication of microbial reduction.

Enterococcus faecium log reduction vs. Hunter L-value, by temperature, at 75 mm bed depth and 1.3 m/s air velocity.
Summary and Conclusions
Because of the close correlation of E. faecium and Salmonella in relative heat stability, this study indicated that Salmonella reductions of 5-log CFU/g and higher may be obtained with typical industrial dry roasting parameters for peanuts. The peanuts located in the middle of the roasting bed were shown to have the lowest microbial reduction. Processing parameter changes that allowed the heated air used for roasting to more effectively heat the middle of the bed (longer time, increased airflow or shallower bed depth) improved reductions and reduced variation in reduction among the top, middle and bottom of the bed. Data from the study also suggested that Salmonella reductions correlate to the Hunter L-value of ground, roasted peanuts. This information provides processors practical guidance to better understand and utilize data from microbial validations performed on specific process parameters in their facilities and, as needed, in selection and modification of those parameters to achieve desired Salmonella reductions during industrial roasting of peanuts.
Acknowledgements
The authors would like to thank Kathleen Giedraitis for her invaluable assistance in running the tests, analyzing the data and literature surveys, as well as Steven Hinkle for running the tests and assisting with data analysis.
Disclaimer: Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Literature Cited
Almond Board of California 2007a Guidelines for validation of dry roasting processes http://www.almondboard.com/Handlers/Documents/Dry-Roast-Validation-Guidelines.pdf .
Almond Board of California 2007b Guidelines for process validation using Enterococcus faecium NRRL B-2354 http://www.almondboard.com/Handlers/Documents/Enterococcus-Validation-Guidelines.pdf .
Bari L Nei D Sotome I Nishina I Isobe S and Kawamoto S 2009 Effectiveness of sanitizers, dry heat, hot water, and gas catalytic infrared heat treatments to inactivate Salmonella on almonds Foodborne Pathog. Dis. 6 : 953 – 958 .
Bell C and Kyriakides A 2009 Chapter 18. Salmonella In: Blackburn C. d. W and McClure P.J (eds.) Foodborne Pathogens – Hazards, Risk Analysis and Control. 2nd ed Woodhead Publishing , Oxford, UK .
Beuchat L.R and Mann D.A 2011 Inactivation of Salmonella on pecan nutmeats by hot air treatment and oil roasting J. Food Protect. 74 : 1441 – 1450 .
Beuchat L.R Komitopoulou E Beckers H Bourdichon F Fanning S Joosten H.M and Ter Kuile B.H 2013 Low-water activity foods: increased concern as vehicles of foodborne pathogens J. Food Protect. 76 : 150 – 172 .
Branch W.D 2007b Registration of ‘Georgia-06G’ peanut J. Plant Reg. 1 : 120 (doi: 16 10.3198/jpr2006.12.0812crc)
Brandon H 2013 Peanuts: more and better variety options Delta Farm Press , Accessed Oct 2013: http://deltafarmpress.com/peanuts/peanuts-more-and-better-variety-options
Calhoun S Post L Warren B Thompson S and Bontempo A.R 2013 Prevalence and concentration of Salmonella on raw shelled peanuts in the United States J Food Protect. 76 : 575 – 579 .
Chang A.S Sreedharan A and Schneider K.R 2013 Peanut and peanut products: a food safety perspective Food Control. 32 : 296 – 303 .
Danyluk M.D Jones T.M Abd S.J Schlitt-Dittrich F Jacobs M and Harris L.J 2007 Prevalence and amounts of Salmonella found on raw California almonds J. Food Protect. 70 : 820 – 827 .
Du W.-X Abd S.J McCarthy K.L and Harris L.J 2010 Reduction of Salmonella on inoculated almonds exposed to hot oil 73 : 1238 – 1246 .
Farakos S.M Frank J.F and Schaffner D.W 2013 Modeling the influence of temperature, water activity, and water mobility on the presence of Salmonella in low-moisture foods Int. J. Food Micro. 166 : 280 – 299 .
Fennema O.R 1996 Chapter 2. Water & Ice , Pages 17 – 50 In: Feenema O.R (eds.) Food Chemistry 3rd ed Marcel Dekker , New York .
Food and Drug Administration 2009 Guidance for industry: measures to address the risk for contamination by Salmonella species in food containing a peanut derived product as an ingredient, Accessed Oct 2013: http://www.fda.gov/Food/GuidanceRegulation/GuidanceDocumentsRegulatoryInformation/ProducePlantProducts/ucm115386.htm .
Food and Drug Administration 2012 In: Lampel K.A Al-Khaldi S and Cahill S.M (eds.) Bad bug book: foodborne pathogenic microorganisms and natural toxins. 2nd ed Accessed Oct 2013: http://www.fda.gov/downloads/Food/FoodborneIllnessContaminants/UCM297627.pdf
Ma L Zhang G Gerner-Smidt P Mantripragada V Ezeoke I and Doyle M.P 2009 Thermal inactivation of Salmonella in peanut butter J Food Protect. 72 : 1596 – 1601 .
McDaniel K.A White B.L Dean L.L Sanders T.H and Davis J.P 2012 Compositional and mechanical properties of peanuts roasted to equivalent colors using different time/temperature combinations 77 : C1292 – C1298 .
Miksch R.R Leek J Myoda S Truyen N Tenney K Svidenko V Greeson K and Samadpour M 2013 Prevalence and counts of Salmonella and enterohemorrhagic Escherichia coli in raw, shelled runner peanuts J. Food Protect 76 : 1668 – 1675 .
Sanders T.H and Calhoun S 2014 Effect of oil and dry roasting of peanuts on survival of Salmonella and Enterococcus faecium Peanut Sci. 41 : 65 – 71 .
Scallan E Hoekstra R.M Angulo F.J Tauxe R. V Widdowson M-A Roy S.L Jones J.L and Griffin P.M 2011 Foodborne illness acquired in the United States-major pathogens Emerg. Infect. Dis. 17 : 7 – 15 .
Shachar D and Yaron S 2006 Heat tolerance of Salmonella enterica Serovars Agona, Enteritidis, and Typhimurium in peanut butter J. Food Protect. 69 : 2687 – 2691 .
Scharff R.L 2012 Economic burden from health losses due to foodborne illness in the United States J. Food Protec. 75 : 123 – 131 .
Torlak E Sert D and Serin P 2013 Fate of Salmonella during sesame seeds roasting and storage of tahini 163 : 214 – 217 .
Wittenberger K and Dohlman E 2010 Peanut outlook: impacts of the 2008–2009 foodborne illness outbreak linked to Salmonella in peanuts, USDA-ERS Report OCS-10a-01 .
Notes
- Buhler Aeroglide, Cary, NC 27511
- USDA ARS Market Quality & Handling Research Unit, Raleigh, NC 27695
- Dept. of Food, Bioprocessing & Nutrition Sciences, North Carolina State University, Raleigh, NC 27695 * Corresponding author's E-mail: dan.poirier@buhlergroup.com
Author Affiliations