

Introduction
Cultivated peanut (Arachis hypogaea L.) is a self-pollinated allotetraploid (2n = 4x = 40) and is the second most economically important legume in the U.S. Peanut is susceptible to many pathogens, with most damage being caused by fungi (Melouk and Backman, 1995). Soil-borne fungi cause diseases that adversely affect peanut health and productivity throughout the growing areas of the United States. Diseases such as pod rot (Rhizoctonia solani Kühn, Pythium myriotylum), crown rot (Aspergillus niger Teigh) and southern blight (Sclerotium rolfsii Sacc) occur in all U.S. peanut-producing areas, while others such as Sclerotinia blight (Sclerotinia minor Jagger) are limited to certain geographic regions. Sclerotinia minor (S. minor), the fungus that causes Sclerotinia blight, white mold and watery soft rots of vegetables, or lettuce drop, can infect host plants from 21 families, 66 genera, and 94 species (Porter and Melouk, 1997). Sclerotinia blight was first observed on peanut in Argentina in 1922 and by 1960 reports of S. minor causing root and pod rot on peanut were published. Sclerotinia blight is now present in most peanut producing countries of the world but losses are most heavily reported in the U.S. The first reports of S. minor in the U.S. were made in Virginia 1971 and then in North Carolina in 1972 (Porter et al., 1982). Sclerotinia blight is of major concern to peanut producers in the Southwest U.S., Virginia and North Carolina. Depending upon severity of field infestation, yield losses due to Sclerotinia blight are typically 10% but may be as high as 50% (Melouk and Backman, 1995). Expensive fungicide applications throughout the growing season are often required for effective disease management. Host plant resistance would provide the most effective solution to managing Sclerotinia blight and limited progress has been made in the development and release of cultivars with enhanced tolerance to the disease (Smith et al., 1991, 1998; Baring et al, 2006).
Several factors contribute to the lack of available Sclerotinia blight resistant cultivars. The inheritance mechanism of host resistance is not well understood. Wildman et al. (1992) suggested that at least two loci were involved in Sclerotinia blight resistance among genotypes studied. Cytoplasmic factors have also been suggested to be involved in Sclerotinia blight resistance (Coffelt and Porter, 1982). Plant morphology can play an important role in resistance to fungal disease because of the environment required for development and progression (Chappell et al., 1995; Coffelt and Porter, 1982; Coyne et al., 1974; Schwartz et al., 1978). Plant types with a more upright growth habit and open canopy, such as Spanish, appear to be more resistant than those with a dense canopy (such as runner and Virginia types) which allows for temperature reduction and moisture accumulation. However, the mechanism of resistance among Spanish types is not purely morphological since the Spanish cultivars Pronto and Spanco are as susceptible as many runner types, suggesting contribution by a genetic component.
Another limitation for breeding programs developing S. minor resistant cultivars is the lack of known sources of resistance. Cultivated peanut has an extremely narrow genetic base which may have resulted from a single domestication event (Simpson et al., 2001) and subsequent inbreeding among a few select parental lines in commercial breeding programs. The advent of molecular genetics gave peanut breeders new found hope that undiscovered germplasm diversity would soon become apparent and available for use. However, early examinations of the genetic diversity in cultivated peanut revealed little or no polymorphism among the accessions studied (He and Prakash, 1997; Kochert et al, 1991; Stalker and Mozingo, 2001). Techniques such as SSR and microsatellite analysis have uncovered some genetic variability in peanut. For example, Hopkins et al. (1999) used simple sequence repeat (SSR) primers to uncover six polymorphic SSRs in cultivated peanut and were able to differentiate 15 of 19 accessions tested. Since that discovery, the number of SSR markers has increased (He et al, 2005). Chu et al. (2007) converted RFLP markers to sequence characterized amplified region (SCAR) markers so as to develop a PCR-based marker system to screen for nematode resistance in peanut. Markers have also been developed for the high-oleate trait (Chu et al., 2009; Chen et al., 2010).
A continued supply of genetic diversity including new or improved variability for target traits is essential for successful crop improvement. Peanut germplasm collections are the best genetic resources for detailed characterization of important traits and are “gold mines” for analysis of allelic diversity. The efficiency of peanut improvement programs, whether conventional breeding alone or powered with marker assisted selection (MAS), depends on the accurate phenotypic data which can then be correlated with genotypic marker data. Establishing a highly significant genotype–phenotype association is one of the prerequisites for efficient breeding with MAS. Linkages or associations between target traits genes and molecular markers are detected based on genetic linkage or association mapping experiments. Molecular markers associated with phenotypic traits have proven extremely useful in breeding programs, either for the characterization of members of segregating populations or for the selection of desired parental genotypes so as to pyramid desired traits (Chu et al., 2011). Markers can also be used as tools for screening germplasm collections for possible new sources of desired traits to be incorporated into adapted lines. Chenault et al. (2009) used association mapping to identify an SSR marker closely associated with Sclerotinia blight resistance in peanut. Chamberlin et al. (2010) used that molecular marker to screen the U.S. peanut mini core collection. In that study, 96 available accessions were genotyped using the marker and 39 total accessions from Spanish, Valencia, runner market types were identified as potential new sources of resistance and targeted for further evaluation in field. The marker was not found to be associated with Sclerotina blight resistance in accessions with a Virginia market-type.
The International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) has over 14,000 accessions in their peanut germplasm collection from which a core collection (Upadhyaya et al., 2003) and a mini-core collection (Upadhyaya et al., 2002) have been developed. Since its creation the ICRISAT peanut mini core has been characterized for drought resistance (Upadhyaya, 2005) and for high oleic acid content (Mukri et al., 2012). The objective of this study was to characterize available accessions from the ICRISAT mini core collection using the molecular marker associated with Sclerotinia blight resistance.
Materials and Methods
Plant Materials
One hundred twenty-four (124) accessions from the ICRISAT peanut mini core collection were available and obtained from R. Pittman (Plant Genetic Resources Conservation Unit, USDA-ARS, Griffin, GA). Seed from all viable accessions (Table 1) were germinated and grown to maturity under greenhouse conditions. Descriptor information (Table 1) was provided by N. Puppala (personal communication).
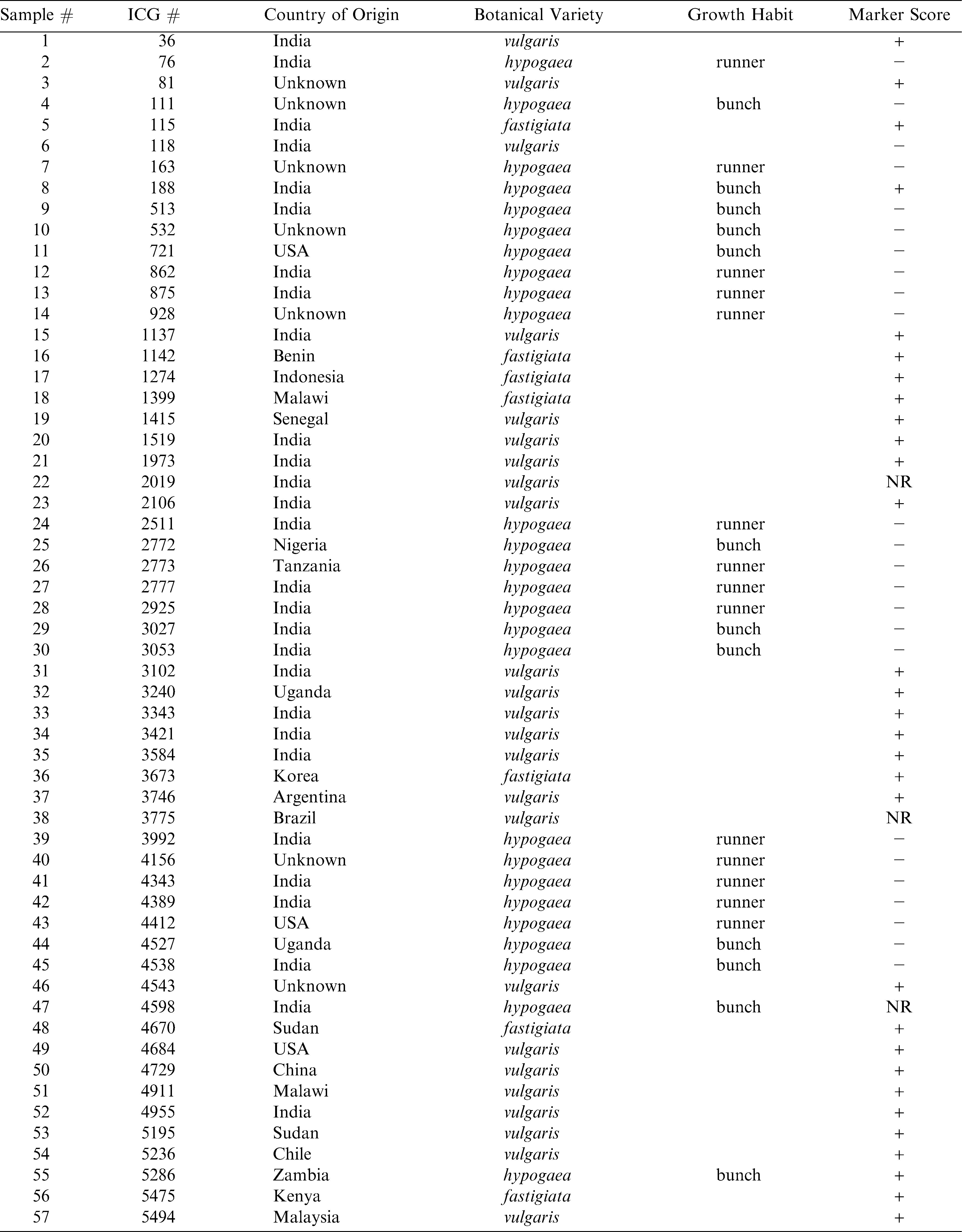
List of available accessions of the ICRISAT peanut mini-core collection included in this study along with available descriptor data and the results of molecular marker analysis.
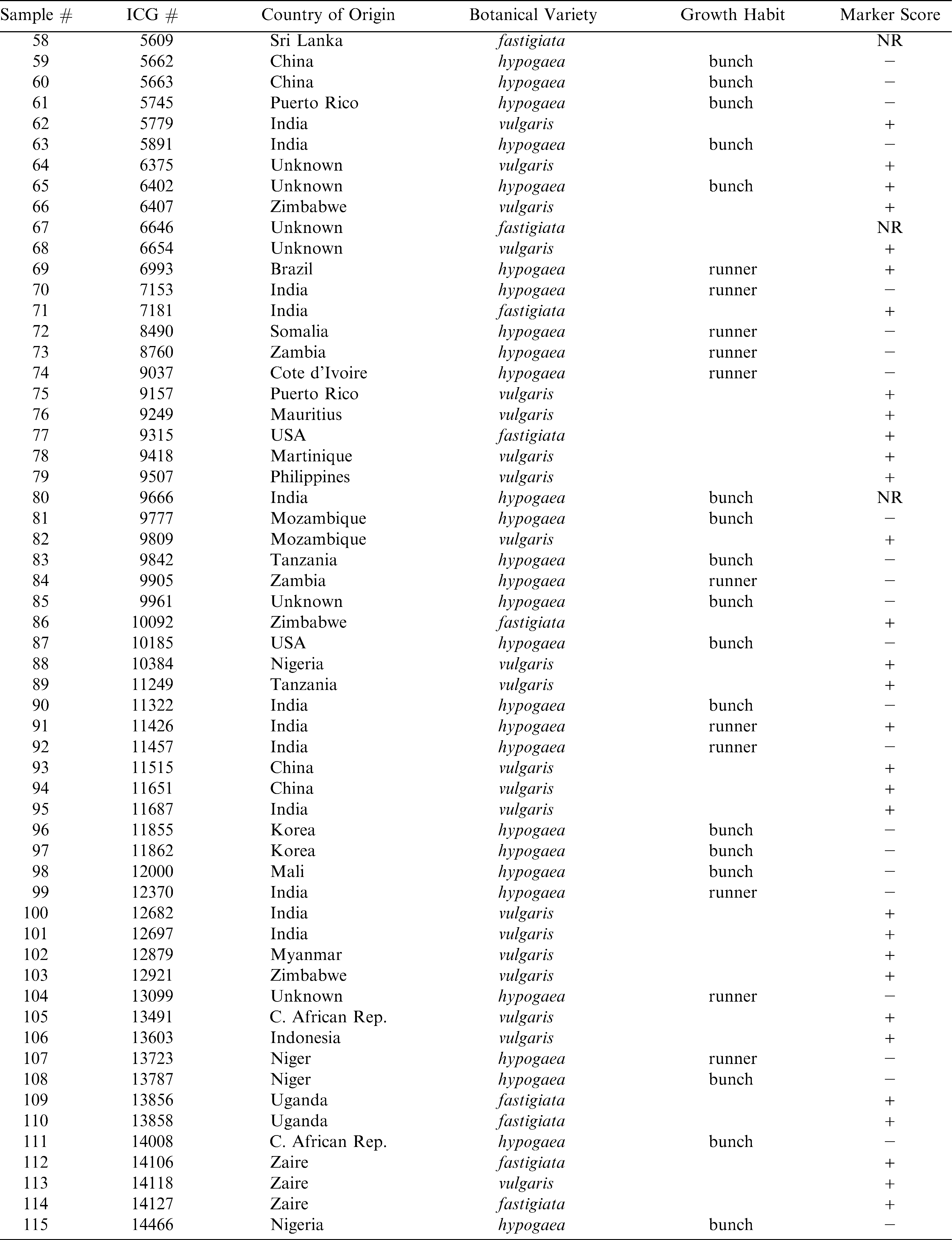
Continued.

Continued.
DNA Extraction and Marker Analysis
DNA was extracted from each genotype listed in Table 1, either from dry, mature seed (Chenault et al., 2007) or from young leaf tissue. In case of the latter, 0.2 g of unfolded leaflet tissue was collected from each plant, de-veined, ground in liquid N2 to a fine powder and vortexed in 1.5 mL extraction mixture [1∶1, extraction buffer (0.1 M Glycine-NaOH, pH 9.0, 50 mM NaCl, 10 mM EDTA, 2% SDS, 1% Na-lauryl sarcosine): phenol-chloroform-isoamyl alcohol (25∶24∶1)]. Extraction mixtures were shaken vigorously for 10 min and then microfuged for 15 min at 10 K rpm at room temperature. DNA was precipitated from the upper layer of each sample by the addition of 750 µL of isopropanol followed by gentle inversion. DNA was spooled onto a glass hook, washed with 70% ethanol, and allowed to air dry for 15 min at room temperature. Hooks were then placed into tubes containing 1 mL extraction buffer and DNA was re-suspended overnight. DNA suspensions were then incubated with 50 µg Proteinase K for 30 min at 37 C. Proteins and other remaining cellular debris were removed by extraction with phenol-chloroform-isoamyl alcohol (25∶24∶1) followed by extraction with ½ volume of chloroform to remove remaining phenol. DNA was precipitated by the addition of 750 µL isopropanol, spooled on glass hooks and allowed to air dry for 1 hr at room temperature. DNA was re-suspended in 100 µL of Tris-EDTA buffer and stored at −20 C until further use.
Amplification using primer pPGPseq2E6L (5′ TACAGCATTGCCTTCTGGTG 3′) and primer Marker 3 (5′ GCACACCATGGCTCAGTTATT 3′) was carried out in a PTC-100 thermal cycler (MJ Research, Watertown, MA) under conditions previously reported (Chenault et al., 2009). Bands were identified using Quantity One software (Biorad). Each banding pattern was verified by repeating reactions in triplicate. Resulting bands were scored as previously reported (Chenault et al., 2009). Genotypes possessing the 145 bp band associated with Sclerotinia blight resistance were given a score of (+). Those genotypes carrying only the 100 bp band were given a (−) rating. Those genotypes possessing neither band were not rated (NR).
Results and Discussion
Screening plant germplasm collections often results in new genetic resources for use in breeding programs. The U.S. peanut germplasm collection has been screened for possible sources of resistance to late leaf spot (Holbrook and Anderson, 1995), Meloidogyne arenaria (Holbrook et al., 2000a, 200b), pepper spot (Damicone et al., 2010) and Sclerotinia blight (Chamberlin et al., 2010; Damicone et al., 2009). The ICRISAT peanut collection has also been explored for sources of resistance to leaf spot (Mehan et al., 1994; Subrahamanyam et al., 1995), rust (Subrahamanyam et al., 1995), aflatoxin accumulation (Mehan, 1989), rosette virus (Subrahmanyam et al., 2001), peanut bud necrosis virus (Subrahmanyam et al., 1985) and insect pests (Wightman and Ranga Rao, 1994). Total oil and protein contents have been explored (Upadhyaya et al., 2001) and oleic acid content has been defined (Mukri et al., 2012), as well as crop growth rate, water use efficiency and assimilate partitioning (Nageswara Rao et al., 1994).
In this study, a molecular marker shown to be associated with resistance to Sclerotinia blight in peanut (Chenault et al., 2009) was used to characterize available accessions from the peanut ICRISAT mini core collection. In a previous report, Chenault et al. (2009) reported this marker to be significantly associated with resistance seen in 39 peanut genotypes examined in field trials over a 9 year period. Subsequently, this marker has been used to screen the U.S. peanut germplasm mini-core collection and identify possible sources of resistance to Sclerotinia blight (Chamberlin et al., 2010) and was found in accessions of the U.S. core collection shown to be highly resistant in field trials (Damicone et al., 2009, data not shown). Using this marker, successful amplification was achieved for all but 7 available ICRISAT accessions. Lack of amplification for those seven accessions appears to be due to the absence of primer binding site(s) since amplification of the same templates using other primer sets and control primers were successful (data not shown). Data typical of such amplifications is shown in Figure 1 and the genotyping result (marker score) for all accessions is listed in Table 1. Available accessions tested from the ICRISAT collection were from 34 different countries. There was no significant correlation between marker presence and country of origin.
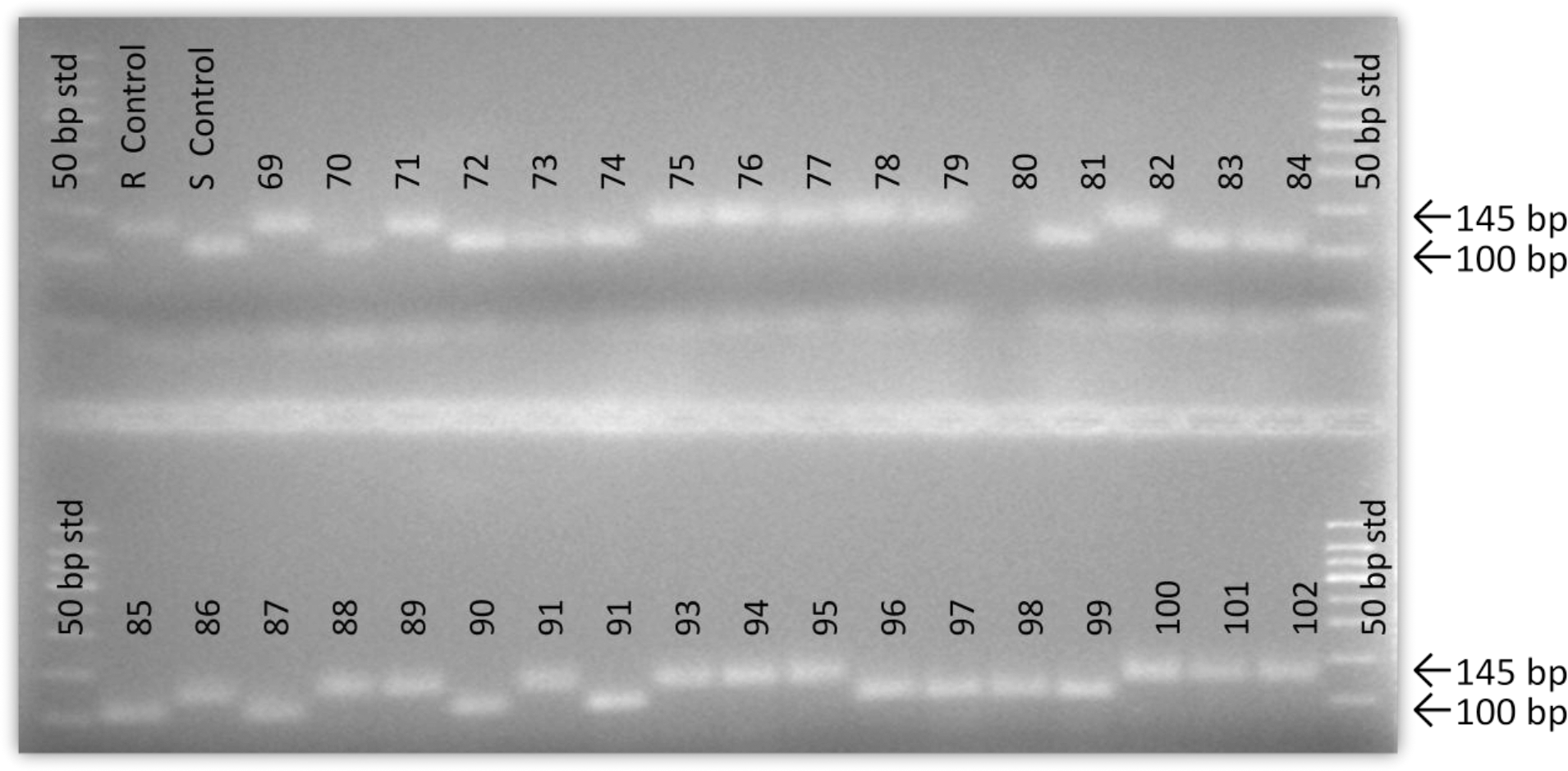
Example of data from SSR genotyping of ICRISAT peanut mini-core accessions. R control = resistant control cv. Red River Runner; S control = susceptible control cv. Okrun.
Categorized by botanical variety, the ICRISAT peanut collection is comprised of 45% var. hypogaea, 35.7% var. vulgaris, 16.1% var. fastigiata, 0.1% var. aequitoriana, 0.14% var. hirsuta, and 1.72% var. peruviana (Upadhyaya et al., 2001). The composition of the group of ICRISAT peanut mini-core accessions available for this study is 47.6% var. hypogaea, 37.1% var. vulgaris and 15.3% var. fastigiata. Genotyping with the SSR marker associated with Sclerotinia blight resistance in peanut reveals that 67/124 (54%) of the accessions were positive for the presence of that marker. Again, classified by botanical variety, 43/46 (93.5%) of var. vulgaris accessions possess the resistance allele, followed by 16/19 (84.2%) of var. fastigiata and 7/59 (11.9%) of var. hypogaea. The fact that the majority (64%) of the accessions classified as var. vulgaris tested positive for the marker is not surprising and is consistent with a previous report of this marker being highly correlated with Sclerotinia blight resistance in Spanish market types (Chenault et al., 2009). It is interesting that, although the marker has also been shown to be tightly linked with resistance in runner market types (var. hypogaea), the majority of hypogaea accessions are not positive for the marker, and of those that do possess the marker, the majority are listed as having a bunch instead of runner growth habit, which is consistent with Virginia market types. This particular SSR marker has not been shown in the past to be correlated with resistance in Virginia market types (Chenault et al., 2009). Of the var. hypogaea accessions that were positive for the marker, three are advanced cultivars, three are landraces, and one is an advanced breeding line. Further investigations with regards to classification of those particular accessions are being conducted to determine if they (1) are true Virginia market types and (2) exhibit elevated levels of resistance to Sclerotinia blight (marker validation). A large majority of accessions classified as var fastigiata (market type Valencia) were positive for the marker (84.2%). The marker has been shown to be consistent with resistance found in Valencia market types (Chenault et al., 2009). Previously, the marker used in this study was shown to be significantly associated with resistance to Sclerotinia blight in peanut cultivars and PIs that had been thoroughly evaluated in field trials (Chenault et al., 2009) and the marker was also used to evaluate the U.S. peanut germplasm mini-core (Chamberlin et al., 2010). Furthermore, the presence of this marker has been confirmed in PIs from the U.S. core germplasm collection that were identified as having enhanced resistance to Sclerotinia blight (Damicone et al., 2009) and those new sources of resistance have been incorporated into peanut breeding programs. There are no reports available where ICRISAT accessions have been phenotyped with regards to Sclerotinia blight resistance, but work is currently underway to achieve that task and further validate the use of this marker for resistance screening.
Summary and Conclusions
In this study, 67 ICRISAT mini-core accessions spanning hypogaea, vulgaris and fastigiata botanical varieties were identified as potential new sources of Sclerotinia blight resistance. Since no reports of field evaluation of the ICRISAT mini-core collection for Sclerotinia blight resistance or susceptibility are available to aid in further validation of this marker as a selection tool for breeding programs, studies are currently underway toward that end. Furthermore, the success of MAS depends upon the establishment of a significant genotype–phenotype association. Although Chenault et al. (2009) established an association of this marker with extensive phenotypic data from cultivar, breeding lines and PIs, more needs to be done to define the extent to which the marker is linked to S. minor resistance and to define the corresponding QTL. Currently, recombinant inbred lines (RILs) are being generated specifically for those genetic linkage and association mapping studies. However, the results obtained in this study have identified accessions worthy of such evaluation in the field for S. minor resistance and also identified those with low probability of being a source of resistance, reducing the amount of field work required to test the ICRISAT peanut mini-core collection by 46%.
Acknowledgements
The author would like to thank Joren Whitley and Kimberly Walker for their technical assistance involved in this research. Appreciation should also be given to Dr. Roy Pittman, former curator of the USDA-ARS peanut germplasm collection located in Griffin, GA, for supplying germplasm accessions for analysis. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Literature Cited
Bannore Y.C Chenault K.D Melouk H.A and El Rassi Z 2008 Capillary electrophoresis of some free fatty acids using partially aqueous electrolyte systems and indirect UV detection. Applications to the analysis of oleic and linoleic acids in peanut breeding lines J. Sep. Sci. 31 : 2667 – 2676 .
Baring M.R Simpson C.E Burow M.D Black M.C Cason J.M Ayers J Lopez Y and Melouk H.A 2006 Registration of ‘Tamnut OL07’ peanut Crop Sci. 46 : 2721 – 2722 .
Chamberlin K.D.C Melouk H.A and Payton M.E 2010 Evaluation of the U.S. Peanut Mini Core Collection Using a Molecular Marker for Resistance to Sclerotinia minor Jagger Euphy. 172 : 109 – 115 .
Chappell G.F Shew B.B Ferguson J.M and Beute M.K 1995 Mechanisms of resistance to Sclerotinia minor in selected peanut genotypes Crop Sci. 35 : 692 – 696 .
Chen Z Wang M.L Barkley N.A and Pittman R.N 2010 A simple allele-specific PCR assay for detecting FAD2 alleles in both A and B genomes of the cultivated peanut for high-oleate trait selection Plant Mol. Biol. Rep. 28 ( 3 ): 542 – 548 .
Chenault K.D Gallo M Seib J.C and James V.A 2007 A Non-destructive seed sampling method for PCR-based analyses in marker-assisted selection and transgene screening Peanut Sci. 34 : 38 – 43 .
Chenault K.D Maas A.L Damicone J.P Payton M.E and Melouk H.A 2009 Discovery and characterization of a molecular marker for Sclerotinia minor (Jagger) resistance in peanut Euphy. 166 : 357 – 365 .
Chu Y Holbrook C.C Timper P and Ozais-Akins P 2007 Development of a PCR-based molecular marker to select for nematode resistance in peanut Crop Sci. 47 : 841 – 847 .
Chu Y Holbrook C.C and Ozias-Akins P 2009 Two alleles of ahFAD2B control the high oleic acid trait in cultivated peanut Crop Sci. 49 ( 6 ): 2029 – 2036 .
Chu Y Wu C.L Holbrook C.C Tillman B.L Person G and Ozias-Akins P 2011 Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut Plant Gen. 4 ( 2 ): 110 – 117 .
Coffelt T.A and Porter D.M 1982 Screening peanuts for resistance to Sclerotinia blight Plant Dis. 71 : 811 – 815 .
Coyne D.P Steadman J.R and Anderson F.N 1974 Effect of modified plant architecture of great northern dry bean varieties (Phaseolus vulgaris) on white old severity, and components of yield Plant Dis. Rep. 58 : 379 – 382 .
Damicone J.P Holbrook C.C Smith D.L Melouk H.A and Chenault K.D 2009 Reaction of the core collection of peanut germplasm to Sclerotinia blight and pepper spot Peanut Sci. 37 ( 1 ): 1 – 11 .
He G and Prakash C.S 1997 Identification of polymorphic DNA markers in cultivated peanut (Arachis hypogaea L.) Euphy. 7 : 143 – 149 .
He G Ronghua M Gao H Guo B Gao G Newman M Pittman R.N and Prakash C.S 2005 Simple sequence repeat markers for botanical varieties of cultivated peanut (Arachis hypogaea L.) Euphy. 142 : 131 – 136 .
Holbrook C.C and Anderson W.F 1995 Evaluation of a core collection to identify resistance to late leaf spot in peanut Crop Sci. 35 ( 6 ): 1700 – 1702 .
Holbrook C.C Stephenson M.G and Anderson A.W 2000a Level and geographical distribution of resistance to Meloidogyne arenaria in the U.S. peanut germplasm collection Crop Sci. 40 ( 4 ): 1168 – 1171 .
Holbrook C.C Timper P and Xue H.Q 2000b Evaluation of the core collection approach for identifying resistance to Meloidogyne arenaria in peanut Crop Sci. 40 ( 4 ): 1172 – 1175 .
Hopkins M.S Casa A.M Wang T Mitchell S.E Dean R.E Kochert G.D and Kresovich S 1999 Discovery and characterization of polymorphic simple sequence repeats (SSRs) in cultivated peanut (Arachis hypogaea L.) Crop Sci. 39 : 1243 – 1247 .
Kochert G Halward T.M Branch W.D and Simpson C.E 1991 RFLP variability in peanut cultivars and wild species Theor. Appl. Genet. 81 : 565 – 570 .
Kottapalli K.R Burow M.D Burow G Burke J and Puppala N 2007 Molecular characterization of the US peanut mini core collection using microsatellite markers Crop Sci. 47 : 1718 – 1727.
Mehan V.K 1989 Screening of groundnuts for resistance to seed invasion by Aspergillus flavus and to aflatoxin production . pp. 323 – 334 In Aflatoxin contamination of groundnut: Proc. Int. Workshop, 6–9 Oct. 1989. ICRISAT Center, India. Patancheru, A.P. 502 324 India.
Mehan V.K Reddy P.M Vidyasagar Rao K and McDonald D 1994 Components of rust resistance in peanut genotypes Phytopath. 84 : 1421 – 1426 .
Melouk H.A and Backman P.A 1995 . pp. 75 – 85 In: Melouk H.A and Shokes F.M (eds.) Peanut Health Management APS Press , St. Paul, Minnesota .
Mukri G Nadaf H.L Bhat R.S Gowda M.V.C and Upadhyaya H.D 2012 Phenotypic and molecular dissection of ICRISAT mini core collection of peanut (Arachis hypogaea L.) for high oleic acid Plant Breed. 131 : 418 – 422 .
Nageswara Rao R.C Singh A.K Reddy L.J and Nigam S.N 1994 Prospects of utilization of genotypic variability for yield improvement in groundnut J. Oilseeds Res. 11 : 259 – 268 .
Porter D.M and Melouk H.A 1997 . pp. 34 – 36 In: Kokalis-Burelle N Porter D.M Rodríguez-Kábana R Smith D.H and Subrahmanyam P (eds.) Compendium of Peanut Diseases, Second Edition APS Press , St. Paul, Minnesota .
Porter D.M Smith D.H and Rodriguez-Kabana R 1982 Peanut Plant Diseases . pp. 326 – 410 In: Peanut Science and Technology , Pattee H.E and Young C.T (eds.) American Peanut Research and Education Society, Inc. , Yoakman, Texas .
Schwartz H.F Steadman J.R and Coyne D.P 1978 Influence of Phaseolus vulgaris blossoming characteristics and canopy structure upon reaction to Sclerotinia sclerotiorum Phytopath. 68 : 465 – 470 .
Simpson C.E Krapovickas A and Valls J.M 2001 History of Arachis including evidence of A. hypogaea progenitors Peanut Sci. 28 : 79 – 81 .
Smith O.D Simpson C.E Grichar W.J and Melouk H.A 1991 Registration of ‘Tamspan 90’ peanut Crop Sci. 31 : 1711 .
Smith O.D Simpson C.E Black M.C and Besler B.A 1998 Registration of ‘Tamrun 96’ peanut Crop Sci. 38 : 1403 .
Stalker H.T and Mozingo L.G 2001 Molecular markers of Arachis and marker-assisted selection Peanut Sci. 28 : 117 – 123 .
Subrahmanyam P McDonald D Waliyar F Reddy L.J Nigam S.N Gibbons R.W Ramanatha Rao V Singh A.K Pande S Reddy P.M and Subba Rao P.V 1995 Screening methods and sources of resistance to rust and late leaf spot of groundnut ICRISAT Info. Bull. No. 47, ICRISAT Patancheru, India .
Subrahmanyam P Naidu R.A Reddy L.J Lava Kumar P and Ferguson M.E 2001 Resistance to groundnut rosette disease in wild Arachis species Ann. Appl. Biol. 139 : 45 – 50 .
Upadhyaya H.D Bramel P.J Ortiz R and Singh S 2002 Developing a mini-core of peanut for utilization of genetic resources Crop Sci. 42 : 2150 – 2156 .
Upadhyaya H.D Ortiz R Bramel P.J and Singh S 2003 Development of a groundnut core collection using taxonomical, geographical and morphological descriptors Genet. Resour. Crop Evol. 50 ( 2 ): 139 – 148 .
Upadhyaya H.D 2005 Variability for drought resistance related traits in the mini core collection of peanut Crop Sci. 45 : 1432 – 1440 .
Wildman L.G Smith O.D Simpson C.E and Taber R.A 1992 Inheritance of resistance to Sclerotinia minor in selected Spanish peanut crosses Peanut Sci. 19 : 31 – 34 .
Wightman J.A and Ranga Rao G.V 1994 Groundnut pests, pp. 395 – 469 In: Smartt J (ed.) The groundnut crop: A scientific basis for improvement Chapman and Hall , London .
Notes
- USDA-ARS, 1301 N. Western, Stillwater, OK 74075 * Corresponding author's E-mail: Kelly.chamberlin@ars.usda.gov
Author Affiliations