

Introduction
Advancements in organic production practices for peanut (Arachis hypogaea L.) have been made in the southeastern U.S., particularly for weed (Johnson et al., 2012; Wann et al., 2011b) and mid-season foliar disease control (Cantonwine et al., 2008; Wann et al., 2011a), but there are still serious obstacles to overcome. One of the key research needs is to improve plant stand without the use of synthetic seed treatments. Untreated peanut seed can result in 50% fewer established plants than seed treated with synthetic seed treatment (Melouk and Backman, 1995). Acceptable plant stands not only maximize yield potential but also minimize pest problems by reducing incidence of tomato spotted wilt virus (Branch et al., 2003), and competition by weeds (Place et al., 2010). Plant stands of around 11 to 12 plants/m of row maximized single row peanut production in conventional management in Georgia (Tubbs et al., 2011), and yields were equal with plant stands of 9 to 11 plants/m using various weed control regimes with organic management in Georgia (Wann et al., 2011b). Weed competition is a significant concern for organic growers since there are fewer options to control weeds in organic production systems than conventional systems.
There are several seedborne and soilborne pathogens that can attack peanut seed or seedlings, including Rhizopus spp., Penicillium spp., Fusarium spp., Aspergillus niger, and A. flavus (Sullivan, 1984). Rhizopus seed rot typically occurs during imbibition or just after germination, and is most common in cool-wet planting conditions (Melouk and Backman, 1995; Sullivan, 1984). Aspergillus crown rot, caused by A. niger, can attack the plant prior to emergence or after emergence before stems have hardened off significantly, and is more common in hot-dry conditions (Jackson and Bell, 1969; Melouk and Backman, 1995). A study conducted in North Carolina evaluating the efficacy of peanut seed or soil treated with organically acceptable seedling disease treatments, such as biocontrol organisms, activated charcoal, copper hydroxide, or hot water showed that seed treated with the biocontrol agent Bacillus subtilis showed the most promise among the other treatments tested (Ruark and Shew, 2010). Cantonwine et al. (2011) found that genotype and seed integrity were more important factors than B. subtilis seed treatment to improve plant stand and reduce Aspergillus crown rot, but the rate of the B. subltilis seed treatment used in the experiment was lower than that used by Ruark and Shew (2010).
The objectives of this research were to assess the direct toxicity of various seed treatments against Rhizopus and Aspergillus species, and relate these findings to the efficacy of these materials for improving plant stands and reducing postemergence plant mortality (damping-off) (Fig. 1). Effects on yield and time required for hand weeding were also investigated.

Peanut plants with symptomology of plant mortality (damping-off). Brown necrotic plant that emerged and died (far left); green but folded leaves from stressed and dying plant (center); and healthy fully expanded leaves from healthy plant (far right).
Materials and Methods
Laboratory Assay
Media-based assays were conducted in the laboratory at Valdosta State University from 2008 to 2012 to evaluate the potential of organically acceptable seed treatments to suppress growth of Aspergillus species or Rhizopus on peanut seed. Experimental units consisted of 100 mm diameter potato dextrose agar (PDA) plates with four peanut seed (cv. Tifguard [Holbrook et al., 2008]). Seed treatments included 1) untreated, 2) conventional (Dynasty [azoxystrobin + fludioxonil + mefenoxam], Syngenta Crop Protection, Greensboro, NC), 3) copper sulfate applied dry, 4) copper sulfate plus water, 5) copper sulfate plus cola, 6) Bacillus subtilis (Kodiak or Kodiak HB; Bayer CropScience, Research Triangle Park, NC), and 7) Streptomyces lydicus WYEC-108 (Actinovate STP, Natural Industries, Inc., Houston, TX, or ActinoGrow ST, SipcamAdvan, Research Triangle Park, NC). In the treatments with copper sulfate, the material was ground into a fine powder using a mortar and pestle prior to application to seed, and the water or cola sprays were used as sticking agents. All seed treatments were standardized at 2.5 g product per kg seed. Ten experimental trials were conducted using a randomized complete block design with 6 to 10 replications each. Blocking was done due to differences in light across the lab bench. Two of the trials used seed that were inoculated with spores of R. stolonifer; all others relied on inocula occurring naturally on the seed. Inoculations with R. stolonifer spores were done by tapping a sporulating culture of the fungus into a bag of seed and mixing the seed well to evenly distribute the spores. The experiments were maintained on laboratory benches at room temperature (20–22.5°C). Treatment effects were evaluated as percent seed affected per plate at 3 d for Rhizopus and at 5 and 7 d for Aspergillus. Aspergillus niger and green-spored Aspergillus, presumed to be A. parasitica and A. flavus, were recorded separately. For a seed to be recorded as affected, the fungal colony had to originate at the seed and grow on both seed and agar.
Field Assessment
Field experiments were conducted near Tifton, GA at the University of Georgia (UGA) Lang-Rigdon and Blackshank Farms in 2010 and Ponder Farm in 2011. The trials were conducted on Alapaha loamy sand at Lang-Rigdon, and Tifton loamy sand at Blackshank and Ponder Farms. Peanuts were planted on 27 May 2010, 28 April 2010, and 13 June 2011 at Lang-Rigdon, Blackshank, and Ponder, respectively. In 2010 at both locations, cv. Georgia Green (Branch, 1996) was planted, while Tifguard was planted in 2011. A change in seed was required because Georgia Green seed was no longer available from seed distributors starting in 2011, and Tifguard was identified as a cultivar with very good potential in organic production systems (Wann et al., 2011a). A change in seed genetics likely caused a differentiation in disease resistance, although resistance to A. parasitica and A. flavus are not well known for most cultivars since they are not usually tested for resistance to a pest that is normally easily controlled with fungicidal seed treatments under conventional production conditions, so quantification of this change in resistance potential is not readily available. However, Tifguard had better plant stands than any other tested cultivar in two years of organic management in South Georgia (Wann et al., 2011a), so it was considered the most optimal cultivar of choice for this experiment. All seed used in these trials were Foundation seed with germination exceeding 90 percent. Trials were planted at 20 seed/m of row at approximately 5 cm deep into a conventionally tilled seedbed (moldboard plow followed by rototiller) using a 2-row Monosem precision air planter (Monosem, Inc., Edwardsville, KS). Plots consisted of two rows, spaced 91 cm apart, and were 9.1 m in length at the Lang-Rigdon location, and 7.6 m in length at the Blackshank and Ponder locations. At the Lang-Rigdon and Ponder Farm locations, plots were managed under organically acceptable practices using a flex-tine cultivator (Aerostar, Einböck GmbH & CoKG, Austria) twice per week for the first five weeks after planting for weed control followed by hand weeding by rep on 19–22 July 2010 and 19–22 July plus 10 Aug. 2011. No additional disease or insect control was used. Both of these sites were irrigated to ensure at least 3 cm water per week (irrigation + rainfall). At the Blackshank location, plots were not irrigated, and conventional peanut production practices were used for in-season pest control following UGA Cooperative Extension Service recommendations (Beasley et al., 1997). Peanuts were dug on 15 Oct., 20 Sept., and 14 Nov. followed by harvest on 19 Oct., 24 Sept., and 22 Nov. at Lang-Rigdon, Blackshank, and Ponder locations in their respective years.
A randomized complete block design was used for all three field experiments, using seven, six, and five replications at Lang-Rigdon, Blackshank, and Ponder locations, respectively. The independent variable consisted of five seed treatments including 1) untreated, 2) conventional (Trilex Star [captan + trifloxystrobin + thiophanate-methyl], Bayer CropScience, Research Triangle Park, NC), 3) copper sulfate applied dry, 4) copper sulfate plus cola, and 5) Bacillus subtilis (Kodiak HB). Copper sulfate treatments were ground as described for the lab assay and all materials were applied at 2.5 g product per kg of seed. Stand counts were conducted by counting all emerged seedlings in a 1 m section in six random locations in each plot. These counts were assessed at 1, 2, 3, and 4 wk after planting at the Lang-Rigdon and Ponder locations, and at 2 and 3 wk after planting at Blackshank. In addition, a plant mortality count was conducted by counting all seedlings that had emerged but died. These data were collected at 2, 3, and 4 wk after planting at Lang-Rigdon and Ponder locations, and at 3 wk after planting at Blackshank. The amount of time spent hand weeding was recorded on a per plot basis in both organically managed locations. Yield was collected at all three locations, and grade factors were evaluated for the Lang-Rigdon and Ponder sites.
Data Analysis
For the laboratory assay, records of A. niger and green-spored Aspergillus were pooled for analyses because incidence values of A. niger alone were not high enough to significantly distinguish treatments, and the responses to treatments were similar for all Aspergillus species observed (data not shown). General linear model univariate analysis of variance (IBM SPSS statistics 20, Armonk, NY) was used to analyze percent incidence data, with seed treatment classified as a fixed factor, and trial as a random factor. Tukey's honest significant difference tests (P<0.05) were conducted to distinguish significant differences among seed treatments. For the field experiments, data were statistically analyzed using PROC GLIMMIX in SAS 9.2 software (SAS Institute, 2009), with mean separation by pairwise t-tests. Data were analyzed individually for each experiment instead of combining over year × site locations to account for treatment differences (or lack thereof) observed in the experiments. Correlation analyses were performed using PROC CORR in SAS.
Results and Discussion
In the lab assay, three experimental trials with 26 replications total were used to evaluate treatment effects for each pathogen. A total of 10 trials were conducted, however four trials were excluded due to low pathogen pressure (<25% in untreated controls). Trends observed in these trials were consistent with the trials included in the analyses (data not shown). One trial was excluded due to high pressure (100% of untreated seed affected by Rhizopus), which was the result of adding too much inoculum. The trend observed in this trial differed from the trials included in the analysis and is discussed later. Two of the trials used to evaluate Rhizopus did not include all seed treatments. Therefore, only those treatments that were common to all trials were compared.
There were no interactions between seed treatment and trial, and seed treatment significantly affected incidence of Aspergillus and Rhizopus (P<0.01). Compared to the untreated control, Bacillus subtilis and S. lydicus seed treatments suppressed Aspergillus (P<0.01), with B. subtilis completely suppressing pathogen establishment (Table 1). The dry and cola copper sulfate treatments resulted in similar incidences of Aspergillus to the untreated control (P = 0.299 and 0.154, respectively), while incidence was higher than the untreated control when copper sulfate was applied with water (P<0.01) (Table 1). The effectiveness of B. subtilis against Aspergillus is in agreement with other studies (Kimura and Hirano, 1988; Zhang et al., 2008), while the lack of response by copper sulfate was unexpected, since similar copper containing compounds provided fungicidal activities against A. carbonarius in a lab study using grape-like media (Belli et al., 2006). Copper sulfate did suppress the incidence of Rhizopus, with similar levels of suppression to that of Dynasty (P = 0.161) (Table 1). However, the activity of copper sulfate was almost completely overcome when pathogen pressure was high (data not shown), which suggests limitations to this seed treatment. Compared to the untreated control, B. subtilis had no effect on Rhizopus (P = 0.926), while S. lydicus caused higher incidence (P = 0.021) (Table 1). Streptomyces lydicus has been shown to reduce Pythium and Rhizoctonia solani on pea and cotton seed (Yuan and Crawford, 1995). Rhizopus is not listed as target pathogens for S. lydicus (Cao et al., 2010), and may not have activity against this fungus.
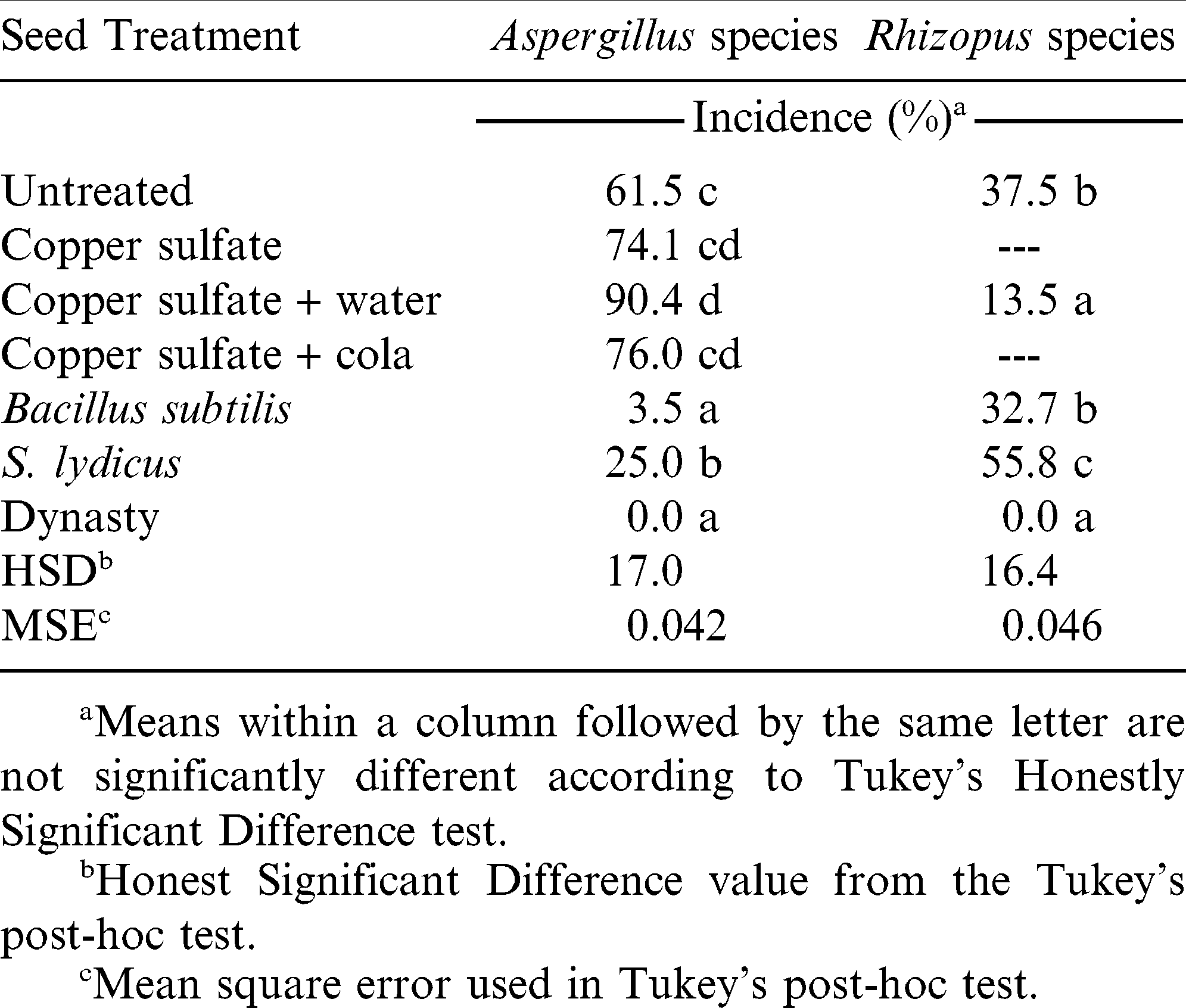
Seed treatment effects on incidence of Aspergillus and Rhizopus in lab assay.
In the field experiments, plant stands were not improved on any sample date with B. subtilis compared to the untreated check (Figs. 2 and 3). There was also equal or greater levels of plant mortality after emergence compared to untreated seed (Figs. 4 and 5), although final totals of both stand counts and dead plants were equal to the untreated in all three field experiments (including the Blackshank location in 2010 – data not shown). Other studies have suggested that B. subtilis has the potential to aid in germination and emergence for improved stand establishment (Cantonwine et al., 2011; Ruark and Shew, 2010; Turner and Backman, 1991). The most favorable responses to B. subtilis have been under stressed conditions, such as seed that were mechanically shelled (Cantonwine et al., 2011), or unfavorable planting conditions from limited water, poor rotation, or cool soils (Turner and Backman, 1991). Yet, in the previous research from South Georgia, only one out of 15 genotypes in each year of the trial had a positive plant stand response that was solely related to B. subtilis seed treatment (Cantonwine et al., 2011). Warm soil temperatures at the 10 cm depth in the Lang-Rigdon-2010 experiment (25 C) and very hot and dry conditions at the Ponder-2011 experiment (36 C) may have caused greater pressure from A. niger to overburden the B. subtilis treatment, or poor establishment conditions for B. subtilis where it could not suppress the pathogen.
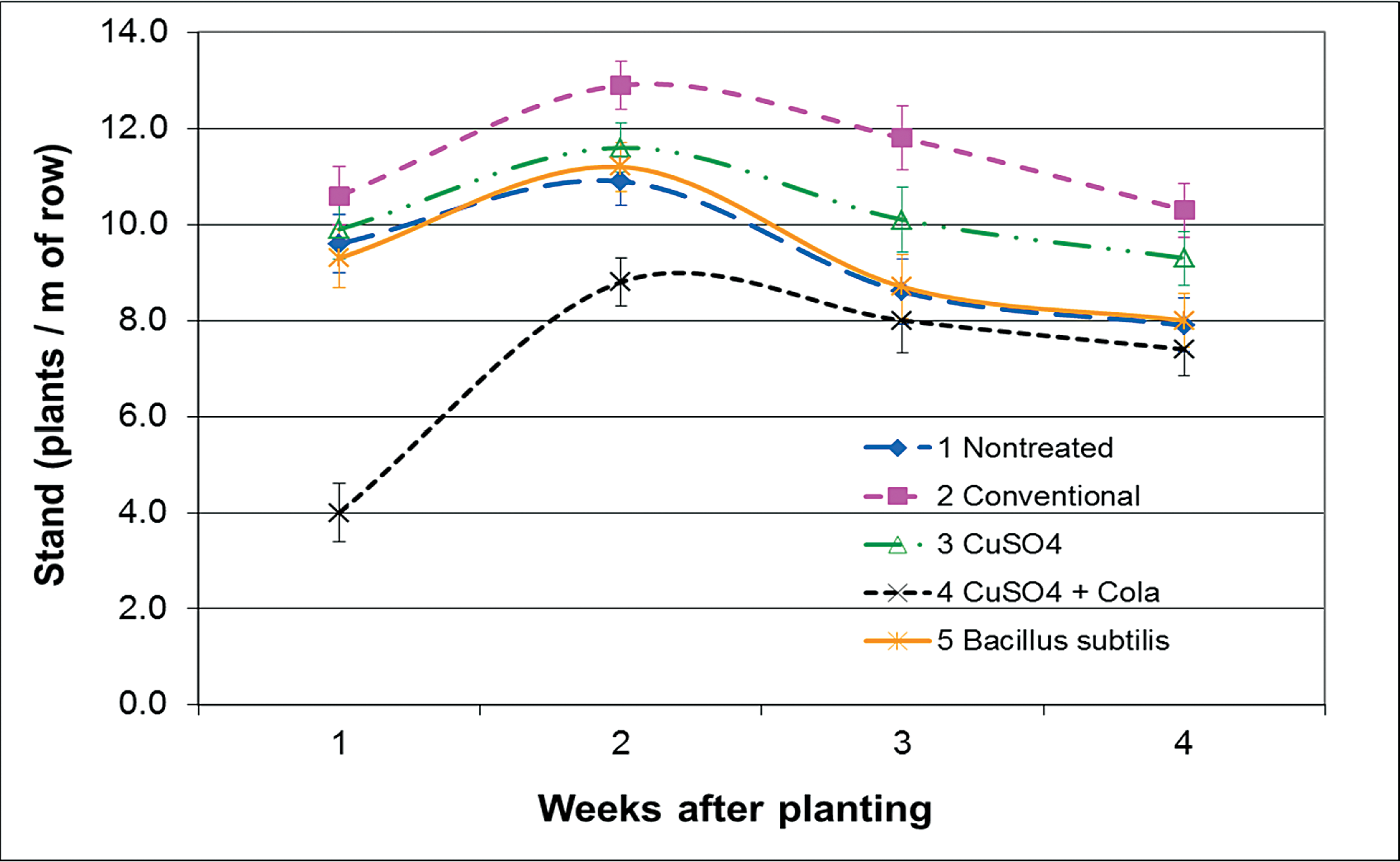
Organic seed treatment plant stands, Lang-Rigdon Farm Tifton, GA, 2010. Peanut planted on 27 May 2010 at 20 seed/m of row. Error bars represent ± standard error of the mean.

Organic seed treatment plant stands, Ponder Farm Tifton, GA, 2011. Peanut planted on 13 June 2011 at 20 seed/m of row. Error bars represent ± standard error of the mean.

Plant mortality counts (seedlings that died after emergence), Lang-Rigdon Farm Tifton, GA, 2010. Peanut planted on 27 May 2010 at 20 seed/m of row. Error bars represent ± standard error of the mean.
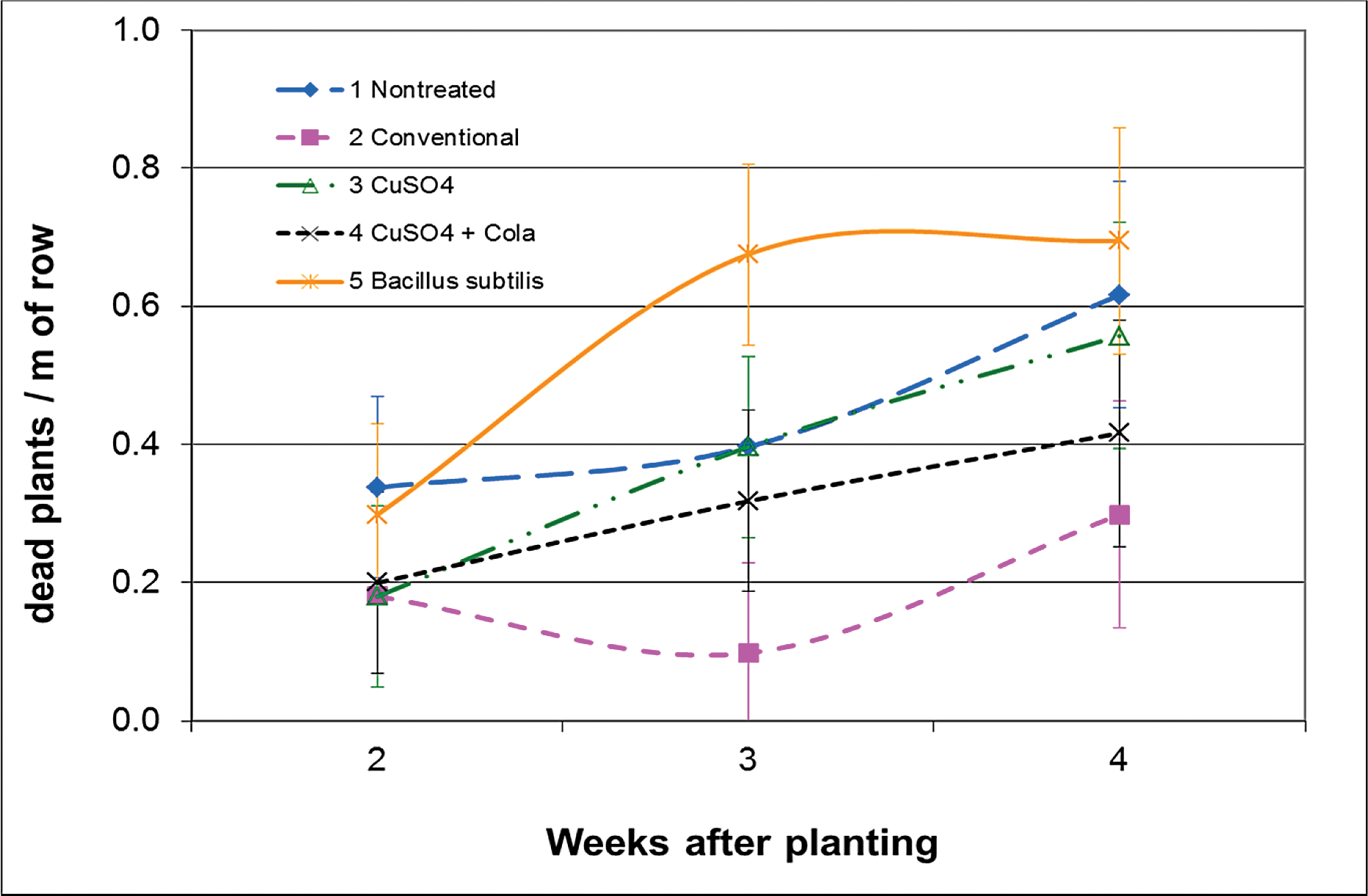
Plant mortality counts (seedlings that died after emergence), Ponder Farm Tifton, GA, 2011. Peanut planted on 13 June 2011 at 20 seed/m of row. Error bars represent ± standard error of the mean.
The only seed treatment that provided some level of efficacy in stand establishment was copper sulfate. When applied dry to the seed, there was an 18% increase in plant stand compared to the untreated seed at 4 wk after planting (Fig. 2), and half as many plants that died after emergence by week 4 at Lang-Rigdon in 2010 (Fig. 4). When copper sulfate was applied after the seed were sprayed with cola, the number of plants that died after emergence was reduced even further, with 79% fewer dead plants than the untreated seed. This is presumably due to greater adherence of the material to the seed. However, emergence of seedlings from the soil was severely delayed compared to all other treatments in both years, and never fully recovered in 2011 at the Ponder location, resulting in a plant stand that was nearly 50% of the untreated seed (Fig. 3). Although care was taken when wetting the seed prior to application of the copper sulfate, it is possible that there was some damage to the seed testae from this process. Such damage is detrimental to seed health, and is one reason the seed industry does not use liquid seed treatments. It is also possible that the additional sugars from the cola attracted facultative pathogens leading to slowed or limited emergence. There was no evidence in the laboratory study of phytotoxicity from a higher rate adherence in the treatments including cola.
Since the lab assays showed good activity from B. subtilis and no benefit from copper against A. niger, yet the field trials displayed no effect from B. subtilis and a positive response where copper sulfate was applied, this would indicate that there were additional variables affecting seedling response to the treatments in this research. It is possible that Rhizopus or other pathogens sensitive to copper sulfate contributed to seedling deterioration during germination, although the hot and relatively dry soil environment in these field studies would have been expected to be very suitable for A. niger. It can also be speculated that there may be some supplemental micronutrient benefit from the copper as well, aiding in growth and a resulting healthier seedling that had greater resistance to damping-off. Peanut seed soaked in low concentrations (10−4M) of copper sulfate, and other inorganic salt solutions, were shown to reduce Aspergillus collar rot (crown rot) in greenhouse and field studies in India (Dasgupta et al., 2000). The authors attributed this effect to enhanced defense responses of the plant rather than direct toxicity to A. niger due to the low concentration of the material. Copper is one of the 13 essential mineral elements needed for plant growth, and copper sulfate is a common micronutrient fertilizer with a nutrient content of 25–35% copper (Brady, 1974). In previous research, copper has increased seed germination for several plant species (Ouzounidou, 1995) and aided in root and shoot growth in alfalfa (Medicago sativa L.) (Peralta et al., 2001), although it can also inhibit germination and growth at high concentrations. The phenomena that caused higher plant stands and reduced mortality with the inclusion of copper in this study was not confirmed; however, it does open opportunities for supplemental research on the effect of copper on germination and stand establishment, and also in reducing damping-off.
Despite variations in plant stand in 2010 at both locations, there were no resulting yield differences (Table 2). However, there were yield differences at the 2011 Ponder location, and this was highly correlated with plant stand (Pearson coefficient = 0.86, P < 0.0001). Plant stand and yield were also highly correlated in another organically managed trial in Georgia (Wann et al., 2011a), which proves the importance of establishing an optimum plant stand in organic production. Although there were no yield differences in 2010, there was a difference in the amount of time required for hand weeding the plots (Table 3). There was likewise a strong correlation between plant stand and time spent hand weeding in the Lang-Rigdon 2010 experiment for all four stand count dates (Pearson coefficient ranging from −0.39 to −0.51, P < 0.025 for all dates). Therefore, whether plant stand affects yield or amount of time spent hand weeding, either way affects net revenue for the organic peanut farmer, such that improved plant stands result in increased income.
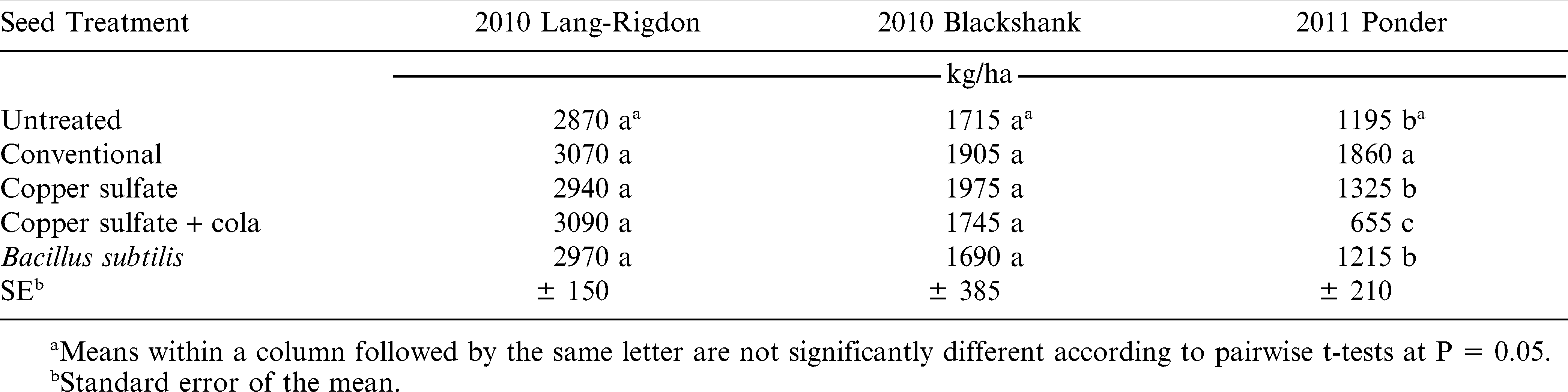
Peanut pod yield under organic management as influenced by seed treatment, Tifton, GA, 2010–2011.
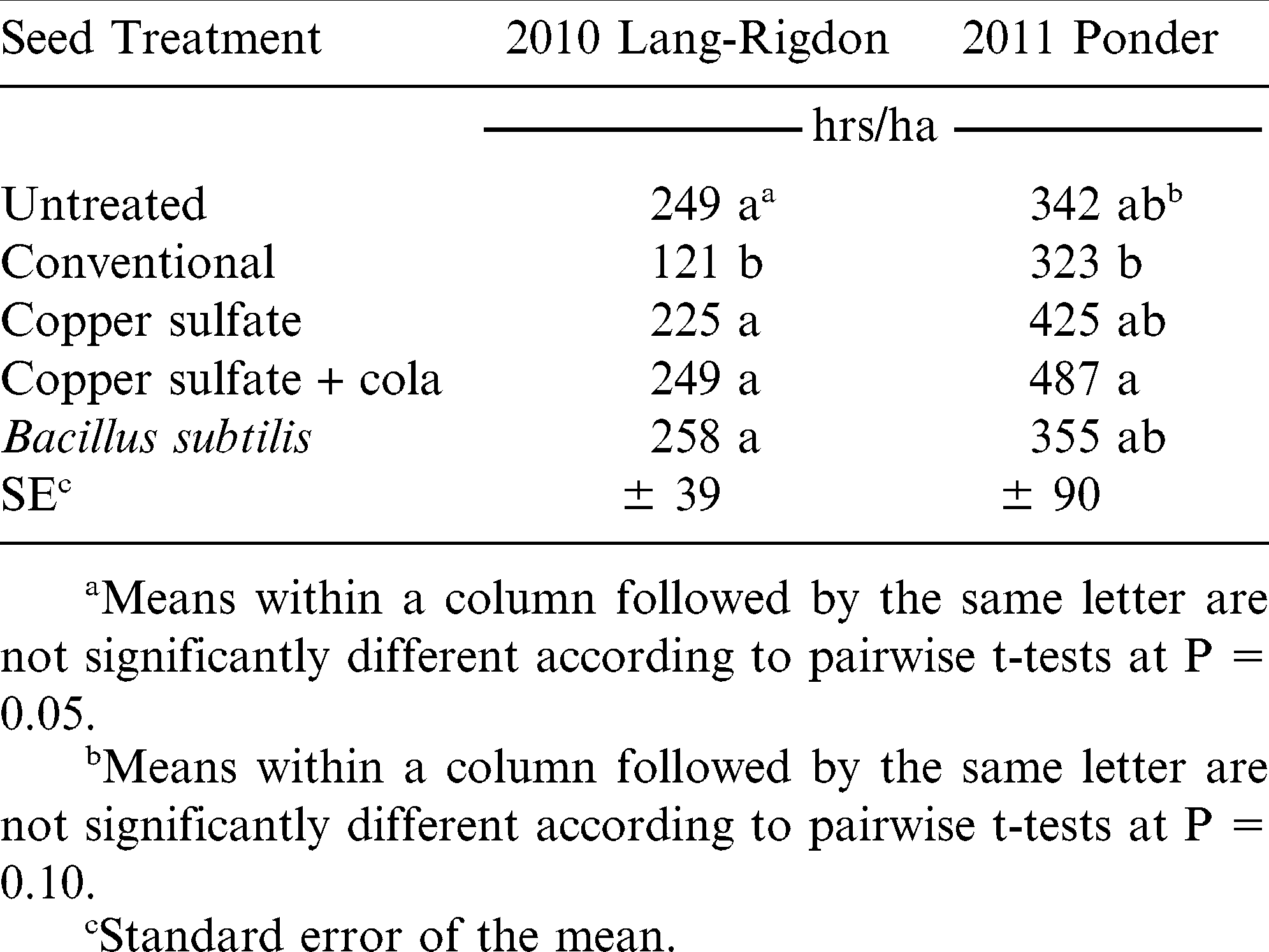
Time requirement for hand weeding plots as influenced by seed treatment, Tifton, GA, 2010–2011.
Summary and Conclusions
Research from the lab assay experiment showed promising results for multiple biological control materials depending on the seed or seedling pathogen in question. However, these effects were not consistent with field results. Based on field results, copper sulfate appeared to have more consistent potential as an organically acceptable material to assist with stand establishment in an organic peanut production system compared to the other materials investigated. The fungicidal activity of B. subtilis against Aspergillus species in vitro did not translate to the field in this study. These data also suggest that copper sulfate may provide an indirect mode of action against the development of Aspergillus crown rot. More research related to this effect would be valuable. Evaluating other particle sizes in combination with an organically acceptable sticking agent that does not damage the seed testae could result in even better activity of copper sulfate on seed and seedling disease.
Acknowledgements
The technical efforts of Chad Abbott, Paige Adams, Jeremy Auerbach, Katie Davis, Hayley Franklin, Rue Hobgood, Colby Johnson, Kati Keeffe, Justin Moss, Jason Sarver, Corey Thompson, Will Vance, Dylan Wann, and Ellie Weeks, plus the statistical expertise and assistance of Jerry W. Davis were essential to the success of this research and are extremely appreciated. We are also grateful to the National Peanut Board and Georgia Peanut Commission for providing financial assistance to make such labor-intensive projects possible.
Literature Cited
Beasley J.P Baldwin J.A Brown S.L Brown S.M Padgett B Bader M.J and Shurley D 1997 Peanut production guide. Agron 95-001 Georgia Coop. Ext. Serv., Univ. of Georgia , Athens .
Belli N Marin S Sanchis V and Ramos A.J 2006 Impact of fungicides on Aspergillus caronarius growth and ochratoxin A production on synthetic grape-like medium and on grapes Food Addit. Contam. 23 : 1021 – 1029 .
Brady N.C 1974 The nature and properties of soils. 8th ed Macmillan Publishing Co., Inc. , New York, NY .
Branch W.D 1996 Registration of ‘Georgia Green’ peanut Crop Sci. 36 : 806 .
Branch W.D Baldwin J.A and Culbreath A.K 2003 Genotype × seeding rate interaction among TSWV-resistant, runner-1 type peanut cultivars Peanut Sci. 30 : 108 – 111 .
Cantonwine E.G Culbreath A.K Shew B.B and Boudreau M.A 2008 Efficacy of organically acceptable fungicides for management of early and late leaf spot diseases on partially resistant peanut cultivars OnlinePlant Health Progress doi:10.1094/PHP-2008-0317-03-RS .
Cantonwine E.G Holbrook C.C Culbreath A.K Tubbs R.S and Boudreau M.A 2011 Genetic and seed treatment effects in organic peanut Peanut Sci. 38 : 115 – 121 .
Cao C Park S and Gardener B.B.M 2010 Biopesticide controls of plant diseases: resources and products for organic farmers in Ohio Fact Sheet. http://ohioline.osu.edu/sag-fact/pdf/0018.pdf .
Dasgupta Raj S.S.K and Srikanta D 2000 Control of collar rot disease of groundnut caused by Aspergillus niger by seed soaking with non-conventional chemicals Indian Phytopath. 53 : 227 – 229 .
Holbrook C.C Timper P Culbreath A.K and Kvien C.K 2008 Registration of ‘Tifguard’ peanut J. Plant Registrations 2 : 92 – 94 .
Jackson C.R and Bell D.K 1969 Diseases of peanut (groundnut) caused by fungi Georgia Agric. Exp. Sta. Bull. 56 .
Johnson W.C Boudreau M.A and Davis J.W 2012 Implements and cultivation frequency to improve in-row weed control in organic peanut production Weed Tech. 26 ( 2 ): 334 – 340 .
Kimura N and Hirano S 1988 Inhibitory strains of Bacillus subtilis for growth and aflatoxin-production of aflatoxigenic fungi Agric. Biol. Chem. 52 : 1173 – 1179 .
Melouk H.A and Backman P.A 1995 Management of soilborne fungal pathogens . pp. 75 – 82 In: Melouk H.A and Shokes F.M (eds.) Peanut Health Management APS Press , St. Paul, MN
Ouzounidou G 1995 Effect of copper on germination and seedling growth of Minuartia, Silene, Alyssum and Thlaspi Biologia Plantarum 37 ( 3 ): 411 – 416 .
Peralta J.R Gardea-Torresdey J.L Tiemann K.J Gomez E Arteaga S Rascon E and Parsons J.G 2001 Uptake and effects of five heavy metals on seed germination and plant growth in alfalfa (Medicago sativa L.) Bull. Environ. Contam. Toxicol. 66 : 727 – 734 .
Place G.T Reberg-Horton S.C and Jordan D.L 2010 Interaction of cultivar, planting pattern, and weed management tactics in peanut Weed Sci. 58 : 442 – 448 .
Ruark S.J and Shew B.B 2010 Evaluation of microbial, botanical, and organic treatments for control of peanut seedling diseases Plant Dis. 94 : 445 – 454 .
SAS Institute 2009 The SAS system for windows. v. 9.2 SAS Inst. , Cary, NC .
Sullivan G.A 1984 Seed and seedling diseases, pp. 37 – 38 In: Porter D.M Smith D.H and Rodriguez-Kabana R (eds.) Compendium of Peanut Diseases / American Phytopathological Society , St. Paul, MN
Tubbs R.S Beasley J.P Culbreath A.K Kemerait R.C Smith N.B and Smith A.R 2011 Row pattern and seeding rate effects on agronomic, disease, and economic factors in large-seeded runner peanut Peanut Sci. 38 : 93 – 100 .
Turner J.T and Backman P.A 1991 Factors relating to peanut yield increases after seed treatment with Bacillus subtilis Plant Dis. 75 : 347 – 353 .
Wann D.Q Tubbs R.S and Culbreath A.K 2011a Genotype and approved fungicide evaluation for reducing leaf spot diseases in organically-managed peanut Online. Plant Health Progress doi:10.1094/PHP-2010-1027-01-RS .
Wann D.Q Tubbs R.S Johnson W.C Smith A.R Smith N.B Culbreath A.K and Davis J.W 2011b Tine cultivation effects on weed control, productivity, and economics of peanut under organic management Peanut Sci. 38 : 101 – 110 .
Yuan W.M and Crawford D.L 1995 Characterization of Streptomyces lydicus WYEC108 as a potential biocontrol agent against fungal root and seed rots Appl. Environ. Microbiol. 61 ( 8 ): 3119 – 3128 .
Zhang T Shi Z.Q Hu L.B Cheng L.G and Wang F 2008 Antifungal compounds from Bacillus subtilis B-FS06 inhibiting the growth of Aspergillus flavus World J. Microbiol. Biotechnol. 24 : 783 – 788 .
Notes
- Dept. of Crop and Soil Sciences, University of Georgia, Coastal Plain Experiment Station, Tifton, GA 31793
- Dept. of Biology, Valdosta State University, Valdosta, GA 31698
- Dept. of Plant Pathology, University of Georgia, Coastal Plain Experiment Station, Tifton, GA 31793 * Corresponding author. E-mail tubbs@uga.edu
Author Affiliations