

Introduction
Peanut (Arachis hypogaea L.) has lagged behind other crop species in the development and use of genetic markers for marker-assisted selection (MAS). Stalker et al. (2009) reviewed the early efforts in advancing the science of genomics in peanut and stressed the need for close cooperation between teams of researchers from multiple disciplines to fully utilize the potential of genomics for peanut improvement. They also pointed out the need for development of appropriate populations that could be used for testing markers to establish associations with traits of interest.
Translation of genomics to breeding requires association of molecular markers with phenotypes and the implementation of cost-effective MAS (Collard and Mackill, 2008). Because of sparse genomic information for peanut, researchers have only been able to take advantage of MAS for a limited number of traits, primarily nematode resistance and high oleic acid (Holbrook et al., 2011). Yet in such a narrowly focused project, tremendous (at least 3-fold) gain in the speed of selection was achieved (Chu et al., 2011). These two traits also were exceptional in that nematode resistance was introgressed from a wild species (Simpson and Starr, 2001) where abundant polymorphisms facilitated discovery of markers (Nagy et al., 2010); and for high oleic acid, genes in fatty acid biosynthesis already had been characterized in other oil seed crops enabling straightforward characterization in peanut and discovery of functional mutations (Jung et al., 2000; Lopez et al., 2002). These two traits also are relatively simply inherited in peanut which is in contrast to many other traits for which breeders must select, e.g. disease resistance, drought tolerance, and yield. With the initiation of the Peanut Genome Project (Anonymous, 2008), the acquisition of genome sequence and potential for discovery of markers will outpace the ability to associate these markers with complex traits that have significant genotype by environment effects, i.e. the bottleneck becomes phenotyping because of the need to collect multiple years of data in multiple locations. Anticipating this need, the peanut breeding community initiated the development of several recombinant inbred line (RIL) populations.
A large group of peanut breeders and molecular geneticists met in the spring of 2008 to develop a plan for identifying molecular markers for MAS. Much of the meeting was devoted to the selection of the most appropriate parents to use for the development of RIL populations that will be needed for mapping and marker development. The group agreed to develop 16 structured RIL populations using a 2 by 8 (common by unique) factorial nested mating association mapping design. Parents were selected to attempt to maximize genetic diversity while meeting practical breeding objectives (Table 1). First, two modern runner cultivars [Tifrunner (Holbrook and Culbreath, 2007), and Florida-07 (Gorbet and Tillman, 2009)] were selected as common parents because runner cultivars account for about 80% of the production in the U.S. Tifrunner was also chosen since according to the Peanut Genome Strategic Plan 2012–2016, it was selected as the candidate genotype for reference genome sequencing and baseline expression profiling by RNA-seq technology. As of January 2013, a rough draft of the peanut genome sequence from Tifrunner has been generated, although considerable refinement and quality control of the assembly must be completed before the sequence is released. Second, the eight unique parents were selected to supply diversity across market classes and botanical varieties and are donors of favorable alleles for enhancing drought tolerance and resistance to most important diseases of peanut in the U.S. The eight unique parents are N08082olJCT (a Bailey derived high oleic breeding line), C76-16, NC 3033 (Beute et al., 1976), SPT 06-06, SSD 6 (PI 576638), OLin (Simpson et al., 2003), New Mexico Valencia A (Hsi and Finker, 1972), and Florunner (Norden et al., 1969).
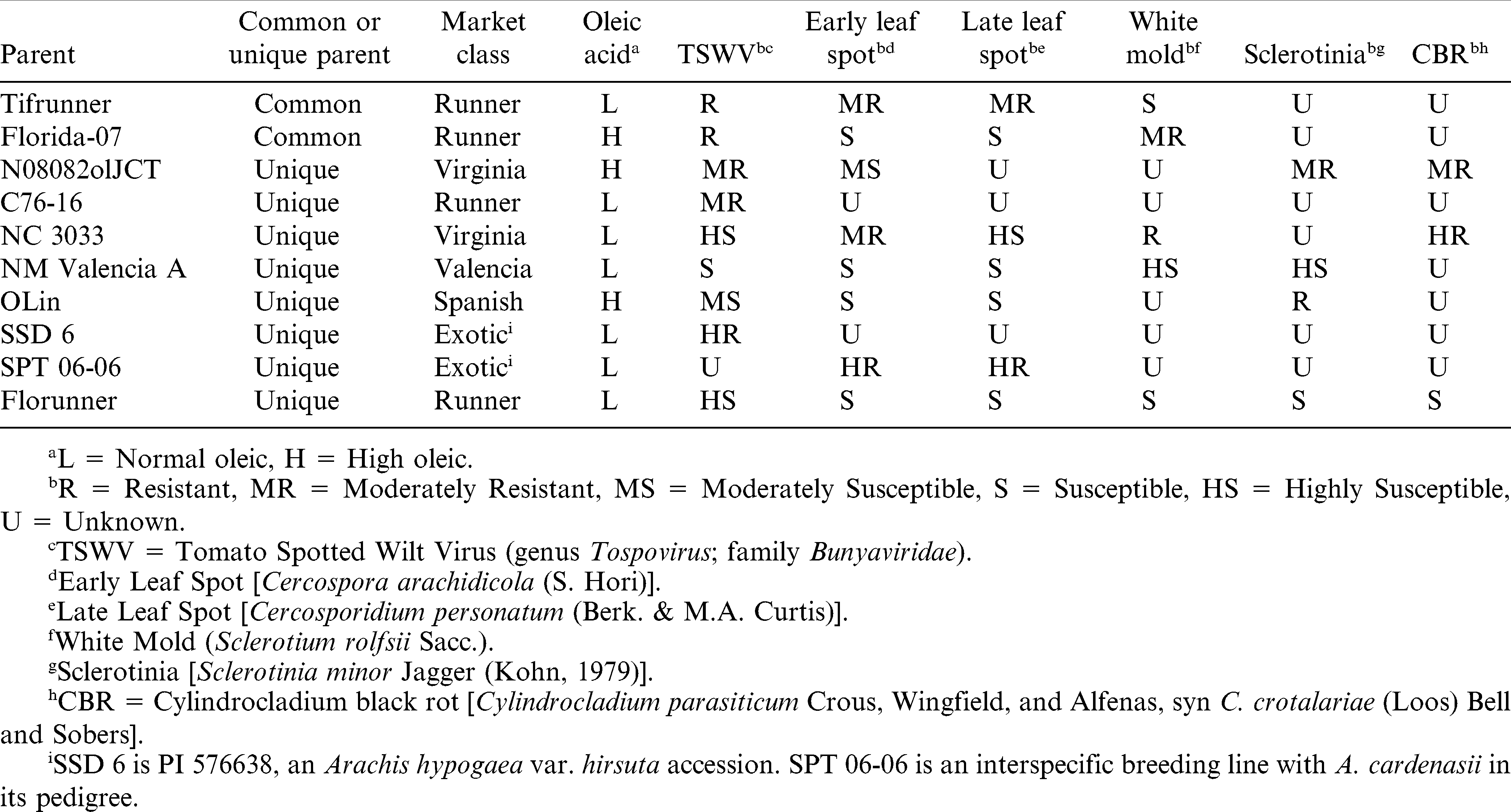
Attributes for parents of 16 structured recombinant line (RIL) populations.
Eight of these cross combinations (Tifrunner × N08082olJCT; Tifrunner × SPT 06-06; Tifrunner × C76-16; Tifrunnner × NC 3033; Florida-07 × N08082olJCT; Florida-07 × SPT 06-06; Florida-07 × C76-16; and Florida-07 × NC 3033) were designated as Set A (Table 2) and were completed in time to send to the 2008-2009 winter nursery. Leaf tissue was sampled from these F1 plants and frozen for DNA extraction and marker analysis to test for hybridity. F2 populations were grown in 2009 in Tifton, GA and Raleigh, NC. Four-hundred F2 plants were harvested from each of the eight populations, and these populations have been advanced using summer and winter nurseries. Generation advance was achieved using small plots of bulked seed to minimize attrition. A random individual plant was harvested from each F6 to provide seed for line increases. Seed increase to provide the community with material for extensive phenotyping of the Set A RIL populations was begun in 2012. F1 seed for the other eight combinations (Tifrunner × SSD 6; Tifrunner × OLin; Tifrunner × New Mexico Valencia A; Tifrunner × Florunner; Florida-07 × OLin; Florida-07 × SSD 6; Florida-07 × New Mexico Valencia A; and Florida-07 × Florunner) designated as Set B, were first sent to the 2009-2010 winter nursery. An update on the current status of these populations is presented in Table 2. Our goal was 400 RILs per population, however, a few populations are slightly smaller than 400 RILs.

Timeline for the development of 16 recombinant inbred line (RIL) populations.
The parents for these populations were selected to sample phenotypic diversity as broadly as possible across elite and exotic gene pools and supply materials for identifying loci underlying the maximum number of traits possible. The segregating populations from these crosses will supply a wealth of material for cultivar development and introgression of novel favorable alleles from elite and exotic sources into modern cultivars. Moreover, segregation of common alleles across populations will increase statistical power for estimating genetic effects necessary for applying MAS in peanut.
It is not feasible to phenotype all populations for all economically significant traits. Input from peanut breeders and molecular geneticists has resulted in a list of populations and traits that should be examined first. Phenotyping projects either underway or proposed with these RIL populations are summarized in Table 3.

Recombinant (RIL) phenotyping in progress and proposed.
Seed increase for most of the RIL populations in set A was begun in the summer of 2012; however, based on expected large variation for resistance to late leaf spot [Cercosporidium personatum (Berk. & M.A. Curtis)], seed increase for the Florida-07 × SPT 06-06 populations was begun in the 2011-2012 winter nursery. This allowed for a late leaf spot phenoytping trial in the field in 2012, and a typical quantitative inheritance was observed in this population. Seventy-nine polymorphic simple sequence repeat (SSR) markers were used to genotype the parents and the 195 RILs included in the late leaf spot field trial, and a low-density map will be constructed in 2013. This population also has potential to be useful for genetic mapping of additional traits such as pod and kernel characteristics and resistance to early leaf spot [Cercospora arachidicola (S. Hori)]. Phenotyping for early leaf spot disease in North Carolina using the genotyped population is planned to begin in 2013.
The ranges of late leaf spot responses in field studies in RIL populations developed from crosses of Tifrunner × GTC-20 and NC 94022 × SunOleic 97R (Qin et al., 2012), and Gregory × Tifguard (Gill et al., 2012) have been large and work is in progress to combine genetic characterization of those populations with disease response data to determine if useful markers for resistance can be developed. Field results from 2012 with the Florida-07 × SPT 06-06 population likewise were promising for late leaf spot, but additional characterization is needed for that population. In addition to field evaluations, Gill et al. (2012) evaluated the Gregory × Tifguard population for components of late leaf spot resistance using lateral stem and detached leaf assays. Studies of this type are time and labor intensive, and are not feasible for large RIL populations. However, component characterization of select lines identified from field studies to represent a range of leaf spot responses should provide valuable information for reference to the genetic profile as well as field disease scores.
The RIL populations with the unique parent NC 3033 crossed with either Florida-07 or Tifrunner are also targeted as high priority populations for phenotyping. The former is expected to segregate for cylindrocladium black rot [Cylindrocladium parasiticum Crous, Wingfield, and Alfenas, syn C. crotalariae (Loos) Bell and Sobers] (CBR) resistance as well as tomato spotted wilt virus (genus Tospovirus; family Bunyaviridae) (TSWV) resistance. The latter should segregate for TSWV, late leaf spot, and white mold (Sclerotium rolfsii Sacc.) resistance, and both should segregate for pod filling percentage.
Research has documented that the parents of the population derived from Tifrunner and NC 3033 diverged substantially for two physiological mechanisms that contribute to drought tolerance (Shekoofa, personal communication). Low hydraulic conductance in the plants appears to be the basis for expression of both traits, and hydraulic conductance is related to the activity of membrane proteins mediating water transport, i.e. aquaporins. The activity of aquaporins can be tested by treating plants with silver, an aquaporin inhibitor (Sadok and Sinclair, 2010). The silver inhibitor test has been effectively applied to a RIL marked population of soybean resulting in the identification of four QTLs (Carpentieri-Pipolo et al., 2012). Phenotyping of the Tifrunner × NC 3033 RIL population using the silver test is expected to lead to QTL markers in peanut.
Previous studies have documented that C76-16 is a unique drought tolerant source for peanut breeding programs (Holbrook et al., 2007; Dang et al., 2012). A pilot study on maturity demonstrated that C76-16 matures significantly earlier than Tifrunner. Phenotyping of the RIL population derived from the cross of Tifrunner × C76-16 should be useful in identifying QTLs underlying drought tolerance and maturity in peanut.
High resolution phenotyping will be essential for associating genetic markers with traits of interest. We have developed an electronic library consisting of more than 40 references on methods to phenotype peanut for economically significant traits. These references are available from the senior author, and should help to standardize the phenotypic data from various locations and projects.
All four populations on which phenotyping is underway will be genotyped with 80–120 SSRs in order to generate a framework map onto which other marker types will be placed. Reduced representation sequencing using a version of RAD-Seq will be carried out initially on parents of all four populations. Bioinformatic analysis of these data will require the development of an analysis pipeline that will distinguish homeologous SNPs from true allelic SNPs since most pipelines already published use relatively short sequences or require a reference genome for alignment, which is not yet available for peanut. Initial analysis of parents, some of which is already underway, along with communication with other testing variations of genotype by sequencing (GBS) for peanut, will provide data for identifying the most cost-effective method for GBS, which we anticipate will be more economical than array-based genotyping. SNP genotyping will be conducted on up to 200 individuals from each population. This will allow the development of a breeder-centric database with many of the basic elements found in Gramene (Ware et al., 2002), Soybase (Grant et al., 2010), and Solanaceae Genomics Network (Mueller et al., 2005), and will be a valuable resource for breeders, molecular biologists, and others engaged in translational genomics and marker-assisted breeding.
Bernardo (2008) developed a thorough and thought provoking perspective article on the lessons learned from 20 yrs of research on QTLs and MAS for complex traits in plants. A major conclusion was that finding QTLs for complex traits is easy and common, but exploiting these QTLs in applied plant breeding programs is much more difficult, and much less common. To increase the probability that discovered QTLs will be used in selection programs, he suggested the active involvement of plant breeders at the beginning of the research project. The selection of parents for the 16 RIL populations for peanut was a multidisciplinary attempt to balance the maximization of genetic diversity while maintaining utility for cultivar development. The continued active involvement of plant breeders, molecular geneticists, pathologists, physiologists, and other disciplines should result in the identification and use of molecular markers for routine development of improved peanut cultivars.
Acknowledgements
The contributions and technical support of Shannon Atkinson, Susan Copeland, Jason Golden, Kathy Marchant, and Betty Tyler are gratefully acknowledged. This work was supported in part by funds from the Peanut Foundation and the National Peanut Board.
Literature Cited
Anonymous 2008 International strategic plan for the peanut genome initiative, Online at http://www.peanutbioscience.com/images/Peanut_Genome_Initiative_StratPlan_Draft_v1.2_Mar08.pdf. Accessed July 18, 2013 .
Bernado R 2008 Molecular markers and selection for complex traits in plants: Learning from the last 20 years Crop Sci. 48 : 1649 – 1664 .
Beute M.K Wynne J.C and Emery D.A 1976 Registration of NC 3033 peanut germplasm (Reg. No. GP 9). Crop Sci. 16 : 887 .
Carpentieri-Pipolo V Pipolo A.E Abdel-Haleem H Boerma H.R and Sinclair T.R 2012 Identification of QTLs associated with limited leaf hydraulic conductance in soybean Euphytica 186 : 679 – 686 .
Chu Y Wu C.L Holbrook C.C Tillman B.L Person G and Ozias-Akins P 2011 Marker-assisted selection to pyramid nematode resistance and high oleic acid trait in peanut The Plant Genome 4 : 110 – 117 .
Collard B.C.Y and Mackill D.J 2008 Marker-assisted selection: An approach for precision plant breeding in the twenty-first century Phil. Trans. Royal Soc. B-Biol. Sci. 363 : 557 – 572 .
Dang P.M Chen C.Y and Holbrook C.C 2012 Identification of genes encoding for drought-induced transcription factors in peanut (Arachis hypogaea L.) Jour. Molec. Biochem. 1 : 196 – 205 .
Gill R Culbreath A.K Holbrook C.C and Ozias-Akins P 2012 Comparison of methods for late leaf spot phenotyping in peanut Proc. Amer. Peanut Res. Educ. Soc. , 44 : 37 – 38 (abstr.)
Gorbet D.W and Tillman B.L 2009 Registration of Florida-07' peanut Jour. Plant Registrations 3 : 14 – 18 .
Grant D Nelson R.T Cannon S.B and Shoemaker R.C 2010 SoyBase, the USDA-ARS soybean genetics and genomics database Nucleic Acids Res. 38 : D843 – D846 .
Holbrook C.C and Culbreath A.K 2007 Registration of Tifrunner peanut Jour. Plant Registrations 1 : 124 .
Holbrook C.C Ozias-Akins P Chu Y and Guo B 2011 Impact of molecular genetic research on peanut cultivar development Agronomy 1 : 3 – 17 .
Holbrook C.C Sullivan D.G Guo B.Z Cantonwine E Wilson D.M and Dong W 2007 Development of peanut germplasm with improved drought tolerance Proc. Amer. Peanut Res. Educ. Soc. 39 : 21 (abstr.)
Hsi D.C.H and Finker R.E 1972 Registration of New Mexico Valencia A peanut1 (Reg No. 14). Crop Sci. 12 : 256 .
Jung S Swift D Sengoku E Patel M Teule R Powell G Moore K and Abbott A.G 2000 The high oleate trait in the cultivated peanut (Arachis hypogaea L.) I. Isolation and characterization of two genes encoding microsomal oleoyl-PC desaturases Mol. Gen. Genet. 263 : 796 – 805 .
Lopez Y Nadaf H.L Smith O.D Simpson C.E and Fritz A.K 2002 Expressed variants of Delta(12)-fatty acid desaturase for the high oleate trait in spanish market-type peanut lines Molec. Breed. 9 : 183 – 190 .
Mueller L.A Solow T.H Taylor N Skwarecki B Buels R Binns J Lin C Wright M.H Ahrens R Wang Y Herbst e.V Keyder R Menda N Zamir D and Tanksley S.D 2005 The SOL genomics network. A comparative resource for Solanaceae biology and beyond Plant Physiology 138 : 1310 – 1317 .
Nagy E.D Chu Y Guo Y Khanal S Tang S Li Y Dong W.B Timper P Taylor C Ozias-Akins P Holbrook C.C Beilinson V Nielsen N.C Stalker H.T and Knapp S.J 2010 Recombination is suppressed in an alien introgression in peanut harboring Rma, a dominant root-knot nematode resistance gene Molec. Breed. 26 : 357 – 370 .
Norden A.J Lipscomb R.W and Carver W.A 1969 Registration of Florunner peanut (Reg No. 2). Crop Sci. 9 : 850 .
Qin H Feng S Chen C Guo Y Knapp S Culbreath A He G Wang M.L Zhang X Holbrook C.C Ozias-Akins P and Guo B 2012 An integrated genetic linkage map of cultivated peanut (Arachis hypogaea L.) constructed from two RIL populations Theor. Appl. Genet. 124 : 653 – 664 .
Sadok W and Sinclair T.R 2010 Genetic variability of transpiration response of soybean [Glycine max (L.) Merr.] shoots to leaf hydraulic conductance inhibitor AgNO3 Crop Sci. 50 : 1423 – 1430 .
Simpson C.E Baring M.R Schubert A.M Melouk H.A Lopez Y and Kirby J.S 2003 Registration of ‘OLin’ peanut Crop Sci. 43 : 1880 – 1881 .
Simpson C.E and Starr J.L 2001 Registration of ‘COAN’ peanut Crop Sci. 41 : 918 .
Stalker H.T Weissinger A.K Milla-Lewis S and Holbrook C.C 2009 Genomics: An evolving science in peanut Peanut Sci. 36 : 2 – 10 .
Ware D Jaiswal P Ni J Pan X Chang K Clark K Teytelman L Schmidt S Zhao W Cartinhour S McCouch S and Stein L 2002 Gramene: a resource for comparative grass genomics Nucleic Acids Res. 30 : 103 – 105 .
Notes
- USDA- Agricultural Research Service, Crop Genetics and Breeding Research Unit, Tifton, GA 31793
- Department of Crop Science, North Carolina State University, , P.O. Box 7629, Raleigh, NC 27695
- Horticulture Department, University of Georgia Tifton Campus, Tifton, GA 31793
- Institute of Plant Breeding, Genetics & Genomics, The University of Georgia, Athens, GA 30605
- North Florida Research and Extension Center, University of Florida, Marianna, FL 32446
- USDA-ARS, Crop Protection and Management Res. Unit, P.O. Box 748, Tifton, GA 31793
- Institute of Plant Breeding, Genetics & Genomics, The University of Georgia Tifton Campus, 31793
- Texas Agricultural Experiment Station, Texas A&M University/Texas Tech University, Lubbock, TX 79401 * Corresponding author (email: Corley.Holbrook@ars.usda.gov).
Author Affiliations