

Introduction
Reducing the period of time required to develop new commercial cultivars is of considerable interest to plant breeders across peanut breeding programs. Accelerated development enables plant breeders to increase the number of breeding generations per calendar year which can considerably improve the efficiency of breeding programs (Sysoeva et al., 2010). The Australian Peanut Genetic Improvement Program (APGIP) has significantly reduced the variety development times, predominately through the use of a winter nursery in coastal north Queensland (latitude 17.1°N) that has enabled two instead of the traditional one generation of peanut to be grown in a 12 month period. Further reductions in variety development times are possible, provided the selection and inbreeding stages can be significantly reduced. Recent work by the wheat breeding team at the University of Queensland has shown that speed breeding techniques can significantly reduce generation times along with concurrent selection for key traits using greenhouse screening methods (Hickey et al., 2009).
Single seed decent (SSD) has been successfully used in international peanut breeding programs, where multiple generations per year have accelerated the inbreeding process to progress fixed lines to multi-site evaluation trials (Holbrook and Culbreath, 2007; Holbrook and Culbreath, 2008; Tillman and Gorbet, 2009). To date, there has been no published work in peanut on the application of recently developed speed breeding technologies used in other crops, which can significantly reduce generation times. Speed breeding technologies involve the growth of plants in controlled environmental conditions (CEnvC), lamps with 24hr high intensity photosynthetic active radiation (PAR), and optimal temperatures (28–32 C) in a greenhouse environment. The only report on the use of 24 hr light growing systems has been that published by the National Aeronautics and Space Administration's (NASA) Advanced Life Support (ALS) program (Rowell et al., 1999). This study concluded that a 24 hr photoperiod resulted in significantly greater vegetative biomass, but lower pod and mature kernel yields compared to the control treatment with a 12 hr photoperiod.
The objective of this study was to assess the potential use of speed breeding techniques in a peanut breeding system. This paper describes a speed breeding system for the rapid development of a population (named cv. P27). This population was inbred from the F2 to F5 generation, with the greenhouse system used for the F2 and F3 generations, and field environments used for F4 and F5 generations (O'Connor, 2012). Only the greenhouse component of this study will be covered in this paper.
Materials and Methods
All experiments described in this paper were carried out in the greenhouse at the Queensland Dep. of Agriculture, Fisheries and Forestry (DAAF) J. Bjelke Petersen Res. Stn. at Kingaroy in south-eastern Queensland (150°50′ E, 26° 33′S).
Preliminary Experiments
Two preliminary trials were conducted to determine the optimal pot culture system to be used in the main trials with the dual objectives of (1) assessing the ideal plant population to be used in large pots and (2) assessing the potential for the 24 hour light system to discriminate genotypes for photoperiod sensitivity. For the preliminary plant population trial, four different grades of seeds were selected from a breeding line (cv. D136-p7-5); with grade 1 (sieve 23), grade 2 (sieve 22), manufacturing grade (sieve 21) and through-sieve oil grade (sieve 20) all sown in 12 × 30 cm pots. Twenty-five seeds of each grade were planted in each pot with three replications. The potting media consisted of two layers, a lower section of two parts krasnozem (sourced from the A horizon of a field trial plot) to one part alluvial sand; and an upper section, 50 cm in depth, of pasteurised peat mix consisting of nine parts alluvial sand, six parts peat moss and one part krasnozem. A granular nitrogen, phosphorus and potassium basal fertilizer was applied to the peat mix prior to sowing, at a rate of 14, three and 10 kg/ha, respectively. Water for the pots was provided manually to the base of each pot as required to maintain pots close to field capacity. Ten days after sowing an emergence count was performed and the pots were then thinned to three different plant populations; 15, 10 and five plants per pot. Each seed grade had one replicate of each plant population. The experiment was carried out in a greenhouse environment under continuous (24 hr) light conditions, with light provided by 450 watt photosynthetically active radiation (PAR) lamps. The PAR lamps were positioned 1 m above the pots with each lamp providing continuous light for six pots. This study was performed in mid to late-winter (sown 26 Jul. 2009), therefore gas heating was provided in the greenhouse and warm water was used for manual irrigation during early stages of growth. The heating regime provided daily maximum temperatures of 28 ± 3C and daily minimum temperatures of 17 ± 3C. Due to the vigorous vegetative growth habit of the plants, pots were rotated twice weekly to ensure any shading effects were minimised. The plants were harvested at 91 days after sowing (DAS) and pod and kernel traits were measured and recorded.
For the preliminary photoperiod trial, three cultivars with suspected differing photoperiod insensitivities were used, Sutherland, a runner type (DPI&F and GRDC, 2006); Wheeler, a virginia type (DPI&F and GRDC, 2003); and TAG-24, an introduced spanish line from India. Ten seeds of each variety were planted in two 30 cm pots, with five seeds per pot. The potting media, fertilizer and watering regimes were the same as used in the plant population trial. The trial was planted on 26 Oct. 2009 and while there was no greenhouse heating required, cooling was supplied through one evaporative cooling unit. Continuous lighting was provided from two 450 watt PAR lamps that were placed 1 m above the pots. These lights operated 24 hr/day to ensure continuous light conditions for the duration of the experiment. Temperature and humidity were variable for the first 15 days after sowing, however for the remainder of the experiment these were kept relatively stable with temperatures at 28 ± 3 C and relative humidities of approximately 60 ± 10%. Ten days after sowing, each pot was thinned to three plants to reduce intra-plant competition. Plants were monitored daily from 25 DAS to determine the onset of flowering. Flowers were tagged and labelled on the day of flowering and inspected daily for successful pegging. After pegging, the tags were transferred from the flowers to the successful pegs and the date recorded. The trial was then grown through to harvest at 100 days after sowing.
F2 and F3 Generation Experiments
The preliminary plant population experiment indicated that 10 plants per pot would be the ideal plant population for a SSD breeding strategy in the 30 cm pots used in the main speed breeding study. The ‘Anova’ pots have previously been used for growing wheat plants under CEnvC and were developed by Ian DeLacy and Mal Hunter at the Univ. of Queensland, School of Land, Crop and Food Sciences (O'Connell, 2007). Forty-one pots were prepared with the same soil media, fertiliser and watering regime as used in the preliminary studies. Four hundred F2 seeds from a cross of one cultivar (Farnsfield) and D147-p3-115, an unreleased breeding line (denoted as cross population P27) were planted randomly on 14 Mar. 2010 into the 41 pots, with 10 parental check plants also included randomly throughout the trial. Seed was planted at a greater rate than 10 seeds per pot to account for less than 100% emergence of seedlings, and later thinned to the 10 plants per pot at 20 days after planting.
Warm greenhouse temperatures during the initial phase of growth necessitated the operation of two evaporative coolers during this early period. Greenhouse daily maximum temperature was maintained at approximately 32 C, with a minimum daily temperature of approximately 22 C. Relative humidity was maintained at approximately 65%, however variability around this average was observed. Due to reduced greenhouse temperature during the first week of April 2010, the gas-fired heating units were operated to maintain an optimum diurnal temperature of 18 ± 3C. The heating units remained in operation for the remainder of the growing period of this F2 generation.
Light was provided by four 450 watt PAR lamps that were suspended one m above the pots (Figure 1). The pots were rotated weekly around the greenhouse benches to ensure all plants received equivalent lighting conditions. At 90 days after planting, a single pot was harvested to assess the maturity of the F3 seed. Harvest observations indicated that the F2 plants were immature and the decision was taken to extend the growing period by a further three weeks. At 113 days, all 41 pots were harvested and a number of pod and kernel characteristics were measured including: number of pods, number of mature pods, pod and kernel weight, number of viable kernels, kernel colour, as well as plant dry weight. After harvest, all kernels were air dried and two viable kernels from each plant were collected and placed in an oven at 38 C for 10 days to break dormancy of the F3 seed.

F2 plants growing in the greenhouse under controlled environment conditions at 15 days after sowing.
The F3 generation was planted on 7 Aug. 2010. Due to a number of plants not producing viable F3 seed, only 270 F3 seeds were sown, in addition to 10 parental checks. A plant population of 10 plants per pot was again used with additional seed planted, where available, to account for emergence losses. Seeds were assigned randomly throughout the pots with all seeds labeled to ensure subsequent generations could be traced back to the individual F2 line. Twenty-eight 30 cm ‘Anova’ pots were used with the same potting media, nutrition, watering regime and continuous light set-up as used for the F2 generation. Gas fired heaters were in use at planting time until the end of September due to cool temperatures experienced during this period. Seedlings were thinned to the required population at 20 DAS. All pots were rotated on a weekly schedule to minimise the effects of shading during the growth period. The harvesting of the F3 generation occurred on schedule at 89 DAS. All 28 pots were harvested and plant, pod and kernel characteristics were measured as for the F2 generation experiment.
Results
Preliminary Experiments
The preliminary plant population trial was characterised by extreme vegetative growth up to harvest at 91 DAS. Despite the vigorous vegetative growth which resulted in intense intra-plant competition within each pot, along with a very early harvest date, most plants produced mature pods and viable kernels. Of the 120 plants harvested, only six plants did not produce a single pod. There were considerable differences in the mean number of pods per plant among the different plant population treatments. The treatment with a population of five plants per pot produced a mean number of 15.8 pods per plant. The ten and fifteen plants per pot populations produced 8.9 and 4.7 pods per plant, respectively (Fig. 2). There was a main effect, predominately linear, of plant population (P< 0.001) and an interaction between population and seed grade (< 0.003), while the main effect of seed grade was not significant (P = 0.46) (Table 1). The standard error of difference between the predicted means of the plant populations was 1.18, with a least significant difference of 2.34.
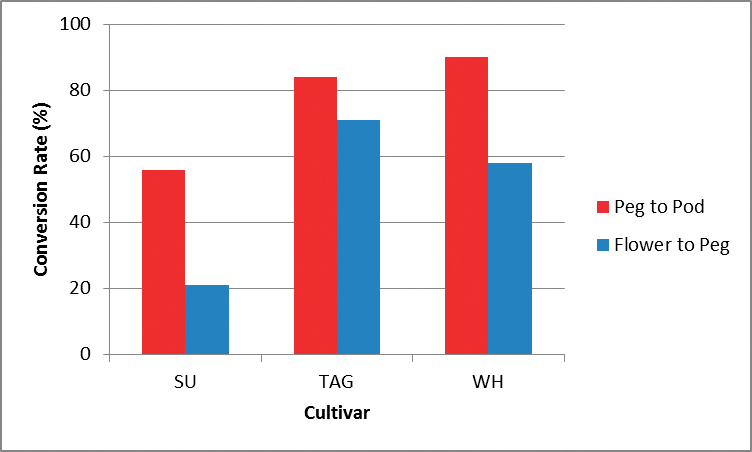
Conversion rate of flower to peg and peg to pod in preliminary photoperiod trial.

Mean number of pods per plant for varying populations and grades.
Within the preliminary photoperiod study, the time to first flower was consistent across the three cultivars, with Sutherland, Wheeler and TAG-24 producing their first flowers at 25, 27 and 25 DAS, respectively. There were however considerable differences between the three cultivars with respect to the success rate of conversion of flowers to pegs. TAG-24 had the highest flower to peg conversion (71%), followed by Wheeler with 58% and Sutherland with 21%, respectively. Sutherland also had fewer flowers during the tagging period with only 42 flowers identified compared to 50 for Wheeler and 52 for TAG-24. There was also a distinct cultivar effect on the conversion of pegs to pods with Sutherland converting 56% of its pegs to pods, compared to 90% for Wheeler and 84% for TAG-24 (Figure 2). This result supports the finding by Nigam et al. (1998) which suggested that pod to peg ratio could be used to indicate genotypic sensitivity to photoperiod.
F2 and F3 Generation Experiments
The use of continuous light in combination with optimum temperature and humidity in the greenhouse facility considerably increased the rate of plant development compared to field conditions. The parental lines, Farnsfield and D147-p3-115, are both full season maturity varieties which require 140 and 145 days in the field to reach full maturity. Using our speed breeding techniques enabled generation time to be reduced to 113 days for the F2 generation and 89 days for the F3 generation. The three week extension required for the F2 growing period was traced back to a decrease in mean temperatures experienced for a 14 day period during May. A mechanical fault in the heating system caused this problem which was rectified immediately after being discovered. During this period there was however a significant decrease in greenhouse temperature, with minimum temperatures decreasing from around 15 C to less than 10 C, as well as a reduction in maximum temperatures form 25 C to 20 C. Temperature was more closely monitored during the F3 generation and no subsequent problems were experienced. There were considerable plant losses observed during both F2 and F3 generations, which resulted from lack of effective emergence and subsequent plant competitiveness in not being able to produce viable pods and/or kernels. From the initial 400 seeds planted in the F2 generation, there was a 68% seed recovery with 270 plants producing viable F3 seeds available for planting. From the F3 seed source, there was a 74% recovery rate, with a total of 201 viable F4 seeds harvested (Table 2). While these plant recovery rates compare favorably with those described by Knauft et al. (1987) for a SSD breeding system, further research is required to determine the actual cause of the plant losses.
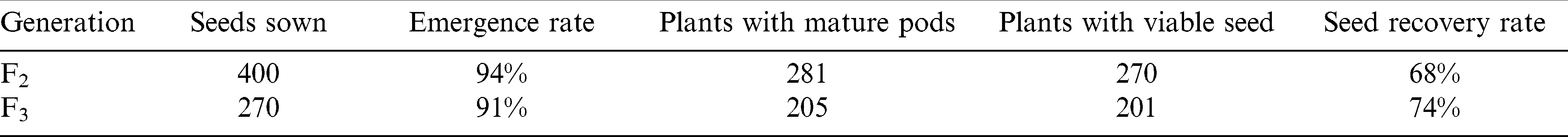
Reproductive success rate of F2 and F3 generations.
Discussion
Overall, the greenhouse speed breeding system was a highly effective strategy for reducing generation times in the peanut breeding program. The controlled environment conditions in conjunction with continuous high intensity PAR light, as described by Hickey et al. (2009), was successfully transferred from a wheat breeding system to peanuts. The system described in this study was able to progress two generations of a full season maturity genotype in 202 days, which is a substantial improvement from the traditional field based pedigree system which would have required around 290 days across two full summer cropping seasons i.e. 17 months in total. The field based system also would have required considerably more land resources for trials and could be more susceptible to adverse environmental conditions during the crop growth cycle. For small-scale breeding programs, limited land, machinery and labour resources are considered to be the main limitations. The greenhouse speed breeding system offers these smaller programs an alternative approach for the rapid advancement of early generation breeding material. We do recognize that this system will require intensive monitoring of both biotic and abiotic stresses. The heater malfunction described in the F2 generation is an example of potential problems that can arise using this system. Severe disease outbreaks have also been reported in the University of Queensland wheat breeding program using controlled environment conditions (Dieters pers comm.), although we have not had any problems in our peanut speed breeding system to date.
There were however considerable challenges in maintaining the CEnvC for the speed breeding system, especially during the cool winter months at our elevated location in Kingaroy in S.E. Queensland, where average minimum temperatures during June, July and August are around 5 C (Bureau of Meteorology, 2012). This is well below the 20 C minimum temperature required for optimal development of peanut plants in the speed breeding system (Ketring, 1982). In the F2 generation experiment, a malfunction in the gas heaters caused a three week delay in the planned harvest date of 90 DAS. Delays such as this can have major implications on planned turnaround for future generations, especially where multi-site field trials are envisaged, as a number of trials must be planted during a relatively short planting window during the November to December period. It is critical that the greenhouse environmental conditions are monitored frequently and that the integrity of heating and cooling systems is assessed on a regular basis. This was done during the F3 generation experiment, where there were no issues with environmental conditions and the trial was harvested as planned at 89 DAS.
The SSD breeding strategy worked well in conjunction with the speed breeding technologies employed during the F2 and F3 generations of this study. As described by Knauft et al. (1987), this breeding method is ideally suited to greenhouse conditions and this proved to be the case in this study. There were, however, plant losses recorded during both greenhouse generations which resulted from either lack of plant vigour following emergence, or plants not being able to produce any pods and/or viable seeds. The recovery rates of between 68% recorded in F2 and 74% in the F3 generations compare favourably to those reported by Knauft et al. (1987), who recommended that SSD breeding programs should assume that ∼70% of seeds planted will produce a plant with at least one seed. It is also possible, however, that the 24 hr light system employed in our study selected out photoperiod-sensitive genotypes, as the preliminary photoperiod experiment strongly indicated that the large genetic variation present for this trait was expressed using the continuous light system. Genetic variation in photoperiod sensitivity has been widely reported in the literature (Bell et al., 1991; Bagnall and King, 1991a; 1991b; Nigam et al., 1994). If proven correct, this could mean that the CEnvC and SSD system employed in this study could be a highly useful technique to easily and effectively select for photoperiodic insensitivity in peanut in early generations.
Indirect support for the above hypothesis comes from the contrasting results observed in the current study compared to the NASA study reported by Rowell et al. (1999). It is hypothesised that the significantly lower seed yield and harvest index observed in the 24 hr light treatment of the NASA study may have been associated with the use of a reputedly highly photoperiodic sensitive cultivar (Georgia Red). Had a photoperiodic insensitive variety been used in this study, seed yield and harvest index may have been similar or even greater under the 24 hr light treatment, as observed for many crop plants when grown under conditions of continuous light (Sysoeva et al., 2010). The positive peanut growth and kernel yield results reported in the current study also suggest that the role of peanuts in the NASA space program should be reinvestigated via the use of photoperiodic insensitive varieties under continuous light. Being a highly nutritious and high oil content food, peanut could offer a highly valuable food source for extended space missions.
Proposed Speed Breeding System to Reduce Variety Development Times in Peanuts
The combination of speed breeding techniques and a single seed decent breeding strategy has the potential to significantly reduce the time in developing new cultivars compared to conventional systems where field based pedigree breeding strategies are commonly employed. Most private and public peanut breeding programs commence preliminary yield (Stage 1) trials by the F5 or F6 generation, by which time the level of heterozygosity has been minimized through the inbreeding process and meaningful selection for more complex quantitative traits made (Isleib et al., 1994; Knauft et al., 1987). Assuming that a breeder starts off with a reasonable quantity of F2 seed (e.g. 500) derived from the selfing of an F1 hybrid plant, and aims to develop fixed F5-derived lines for inclusion in preliminary yield trials, we can conduct a simple analysis of time and cost benefits of using speed breeding and SSD versus field based pedigree breeding approaches.
This analysis also assumes that full season cultivars are being developed, i.e. ones that mature in 145 days using either:
Strategy 1: a pedigree breeding strategy with one summer generation per year
Strategy 2: a pedigree breeding strategy with two generations per year achieved via a summer cycle and a winter nursery in a tropical site capable of producing a second field generation, e.g. near Cairns in North Queensland.
Strategy 3: a speed breeding / SSD breeding strategy using CEnvC and 24 hour lights; and that the F5 generation for all three strategies is grown as spaced plants in the field to generate sufficient seed quantities for inclusion in preliminary yield trials of F5∶6 families.
Figure 3 clearly shows that Strategy 3 (speed breeding / SSD) can reduce the inbreeding development time of the F2 to F5 to around 17 months compared to 42 months using a conventional pedigree breeding approach (Strategy 1). There is also a significant time improvement over Strategy 2 (around 23 months) where a second winter generation is employed (Wright et al., 2011).

Comparison of timeline required to develop F6 fixed lines using Strategy 1, 2 and 3.
The speed breeding system is also less susceptible to adverse biotic and abiotic stresses such as reduced rainfall, low diurnal temperatures and foliar diseases and allows the breeder more flexibility in the generation of new breeding material.
Using speed breeding techniques with continuous light conditions also depends on the cost effectiveness relative to conventional breeding systems (Velez-Ramirez et al., 2011). There are increased monetary costs associated with the speed breeding system, as an analysis of costs of both field based pedigree versus speed breeding / SSD breeding systems have shown (Table 3). Costs were estimated for the range of activities carried out under both breeding approaches used in this study, including planting, harvesting and processing costs in the F2 and F3 generations. In this analysis, it was assumed that the APGIP would develop and grow 25 hybrid populations in the F2 generation in the field, with an average of 400 F2 spaced plants per population. In this pedigree breeding system based on current AGPIP practices where a selection intensity of 10% is applied, around 1000 F2 plants would be selected from a total population of 10,000 F2 plants for further processing (based on visual assessment of plant and pod characters), with around 500 of these plants subsequently selected on the basis of shelling percentage. The seed from these plants are then planted in the field as F2∶3 rows. In the speed breeding / SSD system, it was assumed that 70% of the F2 plants produced a viable seed that was sown in the subsequent F3 generation. There was no selection pressure applied under the SSD system.
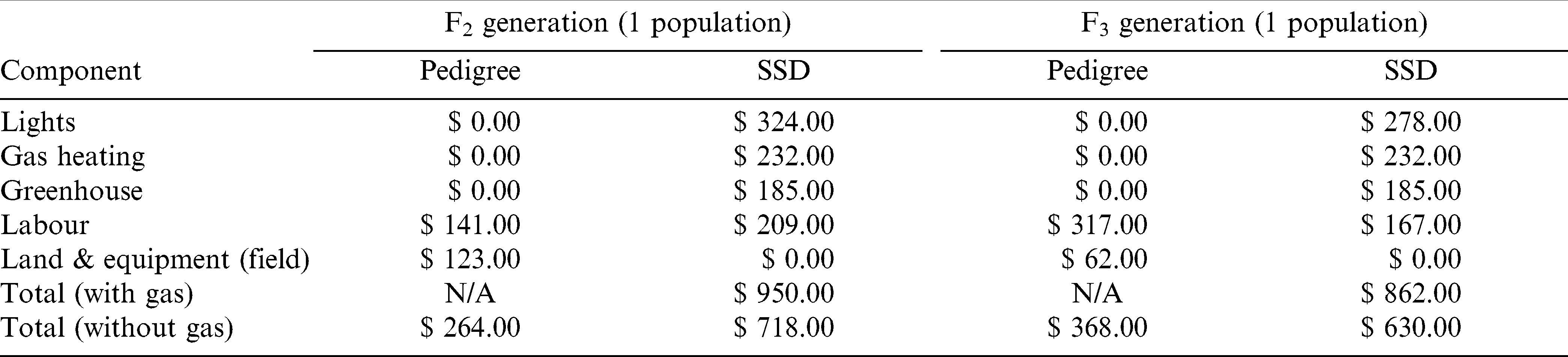
Costs in $US associated with SSD and pedigree systems for one population developed in the F2 and F3 generations.
Cost analysis results expressed on the basis of developing one population in the F2 and F3 generations, indicated the pedigree system was substantially more cost effective in both F2 and F3 generations. With gas heating included, the SSD F2 system costs US$950 per population (US$718 per population without heating) compared to the pedigree system which cost US$264 per F2 population (Table 3). This cost difference between the two systems was reduced to US$494 in the F3 generation due to increased labour costs required in the pedigree system.
Energy (gas and electricity) is a substantial expense for the speed breeding / SSD system, with heating and lighting accounting for around 60% of total cost for generations grown during the winter months. It may be possible to reduce energy usage by using more energy efficient lights and / or converting to a different heating system, e.g. inverter-based air conditioning. Despite the considerable expense of the speed breeding / SSD system there may still be an important role for it to play in the current AGPIP and indeed other global peanut breeding programs, where the universal aim is always to develop better cultivars more quickly.
The above analysis has clearly shown that generation time can be reduced substantially within a speed breeding / SSD system, and hence new cultivars could be developed up to two years quicker compared to using conventional field based pedigree breeding strategies. The increased cost associated with a speed breeding / SSD system may therefore be a relatively small price to pay where more rapid variety commercialization would be able to recover these relatively small upfront costs. Also, this system may be suitable for smaller peanut breeding programs where land and labor resources are limited. Provided the breeder had access to a relatively small greenhouse capable of being fitted out with high intensity lights and heating, a large number of segregating populations, e.g. 25 or more, could still be handled to rapidly reach a level of inbreeding equivalent to that achieved in a field based program.
The speed breeding / SSD system is ideally suited to a backcrossing breeding strategy, where the major objective is to incorporate a relatively simple inherited trait, e.g. one controlled by one or two genes, into a new variety. For example, the high oleic acid trait in peanut, which is controlled by two recessive genes (ol1 and ol2), in runner-type populations, would be an ideal candidate for this strategy. Breeding populations segregating for the high oleic trait can now be rapidly and non-destructively screened using near infrared reflectance (NIR) technology, to facilitate such a back crossing program (Tillman et al., 2006).
Marker-assisted selection (MAS) is becoming widespread in cereal plant breeding programs across the world and has been used successfully to accelerate selection for specific traits. However, collecting reliable phenotypic data for the mass of genomic data generated through next generation sequencing (NGS) technologies is viewed as a roadblock to efficient implementation of MAS (Jackson et al., 2011). The speed breeding system described here is potentially relevant for the rapid development of RIL's which are essential for molecular marker discovery.
Conclusion
The study has demonstrated that speed breeding technologies previously developed for wheat and barley can be successfully transferred to the cultivated peanut and offers peanut breeders a new tool to develop improved cultivars more quickly. The study has clearly shown that generation time can be reduced substantially within a speed breeding / SSD system, and hence new varieties could be developed up to two years quicker compared to using conventional field based pedigree breeding strategies.
Literature Cited
Bagnall D.J and King R.W 1991a Response of peanut (Arachis hypogaea) to temperature, photoperiod and irradiance. 1. Effect on flowering Field Crops Res. 26 : 263 – 277 .
Bagnall D.J and King R.W 1991b Response of peanut (Arachis hypogaea) to temperature, photoperiod and irradiance. 2. Effect of peg and pod development Field Crops Res. 26 : 279 – 293 .
Bell M.J Bagnall D and Harch G 1991 Effects of photperiod on reproductive development of peanut (Arachis hypogaea L.) in a cool subtropical environment. II. Temperature interactions Aus. J. Agric.Res. 42 : 1151 – 61 .
Department of Primary Industries and Fisheries and Grains Research and Development Corporation 2006 Peanut (Arachis hypogaea) Sutherland Plant Varieties J. 20 : 98 .
Department of Primary Industries and Fisheries and Grains Research and Development Corporation 2003 Peanut (Arachis hypogaea) Wheeler Plant Varieties J. 16 : 21 – 22 .
Hickey L.T Dieters M.J DeLacy I.H Kravchuk O.Y Mares D.J and Banks P.M 2009 Grain dormancy in fixed lines of white-grained wheat (Triticum aestivum L.) grown under controlled environmental conditions Euphytica. 168 : 303 – 310 .
Holbrook C.C and Culbreath A.K 2007 Registration of ‘Tifrunner’ Peanut J. Plant Regist. 1 : 124 – 124 .
Holbrook C.C and Culbreath A.K 2008 Registration of ‘Georganic’ peanut J. Plant Regist. 2 : 17 .
Isleib T.G
Wynne J.C
and
Nigam S.N
1994
Groundnut Breeding
. pp.
552
–
623
In
The Groundnut Crop, a scientific basis for improvement
Chapman and Hall
,
London
.
Jackson S.A Iwata A Lee S.H Schmutz J and Shoemaker R 2011 Sequencing crop genomes: approaches and applications New Phytologist. 191 : 915 – 925 .
Ketring D.L
Brown R
Sullivan G
and
Johnson B
1982
Growth physiology
. pp.
441
–
457
In
Peanut Sci. and Technology, Amer
Peanut Sci. Res. and Ed
,
Oklahoma
Knauft D.A
Norden A.J
and
Gorbet D.W
1987
Peanut
. pp.
346
–
385
In
Principles of Cultivar Development. Vol. 2. Crop Species
/
Macmillian Publishing
,
New York
.
Bureau of Meterology 2012 Climate statistics for Australian locations, Bureau of Meterology, Australia, http://www.bom.gov.au/climate/averages/tables/cw_040112.shtml
Nigam S.N Nagweswara Rao R.C and Wynne J.C 1998 Effects if temperature and photoperiod on vegetative and reproductive growth of groundnut (Arachis hypogaea L.) J. Agronomy Crop Sc. 181 : 117 – 124 .
Nigam S.N Nagweswara Rao R.C Wynne J.C Williams J.H Fitzner M and Nagabhushanam G.V.S 1994 Effect and interaction of temperature and photoperiod on growth and partitioning in three groundnut (Arachis hypogaea L.) genotypes Ann. Appl. Biol. 125 : 541 – 552 .
O'Connell L 2007 Speed breeders have they gone potty: or have they found the holy grail Australian Grain. 17 : 40 – 41 .
O'Connor D 2012 Development of rust tolerant peanut genotypes using a single seed descent breeding strategy and speed breeding technologies School of Agriculture and Food Sciences, The Univ. of Qld St Lucia . pp. 1 – 61 .
O'Connor D Wright G.C Baker G Tatnell J Fleischfresser D Dieters M.J and George D 2012 Speeding up release of new peanut cultivars, Proc. of Amer. Peanut Res. and Ed. Soc. 44 : 15 .
Rowell T Mortley D.G Loretan P.A Bonsi C.K and Hill W.A 1999 Continuous daily light period and temperature influence peanut yield in nutrient film technique Crop Sci. 39 : 1111 – 1114 .
Suysoeva M Markovskaya M and Shibaeva T 2010 Plants under continuous light: a review Plant Stress. 4 : 5 – 17 .
Tillman B.L and Gorbet D.W 2009 Registration of ‘AP-4’ Peanut J. Plant Regist. 3 : 138 – 142.
Tillman B.L Gorbet D.W and Person G 2006 Predicting oleic and linoleic acid content of single peanut seeds using near-infrared reflectance spectroscopy Crop Sci. 46 : 2121 – 2126.
Velez-Ramirez A.I van Iepere W Vreugdenhil D and Millenaar F.F 2011 Plants under continuous light Trends in Plant Sci. 16 : 310 – 318 .
Wright G.C Baker G Fleischfresser D and Cruickshank A 2011 Tingoora, a high oleic, ultra early maturing variety bred fro drought and aflatoxin avoidance, Proc. of Amer. Peanut Res. and Ed. Soc. 43 : 79 .
Notes
- Research Scholar, The University of Queensland, School of Agriculture and Food Sciences, St Lucia, QLD, 4072
- Manager (Breeding, Seed and Innovation), Peanut Company of Australia, Haly St, Kingaroy, QLD 4610
- Senior Lecturer, The University of Queensland, School of Agriculture and Food Sciences, St Lucia, QLD, 4072
- Senior Lecturer, The University of Queensland, School of Agriculture and Food Sciences, St Lucia, QLD, 4072
- Honorary Senior Fellow, The University of Queensland, School of Agriculture and Food Sciences, St Lucia, QLD, 4072
- Senior Experimentalist, Department of Agriculture, Fisheries and Forestry, 213 Goodger Rd, Kingaroy, QLD, 4610
- Senior Experimentalist, Department of Agriculture, Fisheries and Forestry, 213 Goodger Rd, Kingaroy, QLD, 4610 * Corresponding author (email: d.oconnor@uq.edu.au)
Author Affiliations