

Introduction
Peanut (Arachis hypogaea L.) is an annual crop of indeterminate growth habit, whose subspecies hypogaea (virginia and runner market types) and fastigiata (valencia and spanish market types) are the most important in crop production (Stalker, 1997). Plant size, presence of floral axes on the main stem (n) and their allocation on branches (n+1, n+2, and n+3) set up the branching pattern and are the basis for the classification of subspecies (Sholar et al., 1995).
The hypogaea botanical types are characterized by the absence of floral axes on the main stem, a branching pattern of alternating pairs of reproductive and vegetative axes, and prostrate (in runner genotypes grown in Argentina), intermediate, or erect growth habit. The fastigiata botanical types presents flowering axes on the main stem, a sequential distribution of axes in other branch types, and upright (erect) growth habit.
Both subspecies have primary branches (commonly called n+1) that originate from the main stem (n). They also have two n+1 branches that originate from the axes located at the axils of cotyledonal nodes and are called cotyledonary branches. The vegetative axes on n+1 (cotyledonary and others) branches bear secondary branches (called n+2), which could be divided into cotyledonary n+2 and other n+2. Only the subspecie hypogaea produces a third type of branch, called n+3 (Shashidhar et al., 1986; Stalker, 1997).
Thus, the combination of growth habits and branching patterns produce different modes of pod distribution in the soil, mainly due to the spatial and temporal variation in pegging and subsequent pod development (Giayetto et al., 2007). This also determines differences in the degree of indetermination and in the length of crop growth season between runner and spanish cultivars. Spanish cultivars, with erect growth habit and sequential branching pattern, have a pod distribution usually concentrated near the base of the main stem and an intermediate to short growing season. Runner cultivars, on the other hand, with prostrate growth habits and an alternating branching pattern, have a scattered pod distribution, and a relatively longer growing season (Giambastiani, 2000).
The different branch types have a differential influence on plant yield, as shown by several studies (Shashidhar et al., 1986; Rehman et al., 2001). Choudhari et al., (1985) noted that in 29 bunch peanut genotypes, primary branches contributed with 86 to 91% of the pod number per plant and, in turn, the first four reproductive nodes contributed on average 85% of the pods. Acknowledging the importance of selecting genotypes with more primary branches per plant, Shashidhar et al. (1986) reported that main stems contribute 6.6%, the n+1 branches 85.2%, and n+2 branches 9.4% of total plant yield of 8 spanish-type cultivars. However, the authors noted that the number of mature pods decreased markedly with each successive (chronologically) n+1 branch, emphasizing that the pods on cotyledonary n+2 contributed significantly to the total yield compared with the others n+1 formed in the upper nodes of the main stem, mainly due to the earlier development of the cotyledonary n+2 branches.
According to Giayetto et al. (2007), 73% of the mature pods were developed on n+1 branches (cotyledonary and others), with significant differences between runner and spanish cultivars. These authors highlighted the greater proportion of mature pods per plant in the spanish cultivar, indicating a pod set more concentrated in time and therefore a lower indetermination, compared to runner cultivars.
The differences of the peanut's plant structure (main stem, types and number of branches) and crop growing season refer to the occurrence of different moments (critical periods) for the determination of pod number and pod weight on each structure. In these moments, those differences will produce interactions between the physiological processes (crop growth rate and partitioning coefficient) of different genotypes (Bell et al., 1993), and temperature and solar radiation (Bell and Wright, 1998). Hence, it is necessary to know the ontogenesis of peanut plant structure and its impact on yield component determinations and plant production of different genotypes. The objective of this study was to describe and quantify, during the reproductive growth of peanut cultivars, the determination moments of pod number and weight for each branch type, and to estimate the relationship with weather conditions prevailing during each respective moment (critical period).
Material and Methods
The study was carried out during the growing seasons of 2009/2010 at the Experimental Area of the Faculty of Agriculture and Veterinary of the Universidad Nacional de Río Cuarto, Córdoba(33°07′ S, 64°14′ W and 421 m above sea level), located in the central region of Argentina. The soil is a typic Hapludoll with fine sandy loam texture. During the peanut growing season (October to April) the normal maximum temperature range is 22.1 to 29.1 C (mean = 26.8 C) and the minimum 9.5 to 17.1 C (mean = 14.5 C). The frost-free period is from 12 Sep to 25 May (256 days). The incident radiation varies between a minimum of 8.5 to a maximum of 30.1 MJ/m2/d and the annual-mean-daily is 20.4 MJ/m2/d. The mean rainfall in the growing season is 656.1 mm and the annual mean rainfall 803.1 mm (Seiler et al., 1995). Two peanut cultivars, Utre, spanish-type-peanut and Granoleico, runner–type-peanut, were hand-sown on three planting dates: 8 Oct. (1st), 10 Nov. (2nd) and 9 Dec. (3rd) in 12 m long with spacing of 0.70 m between rows and 0.08 m between plants. The experimental design was a randomized complete block with a split plot arrangement (Di Rienzo et al., 2011). The sowing dates were the main plot and the cultivars were subplots. The crop was grown without water or nutritional deficiencies and with appropriate weed, pest and disease controls measures to minimize their effect.
Daily climatic data (air and soil temperature, rainfall, and incident solar radiation) were obtained from the agro-meteorological station located on the experimental site. Photosynthetically active radiation (PAR) intercepted by the crop was measured at each sowing date using a Line Quantum Sensor (Li-Cor Inc., Lincoln, NE). Cumulative thermal time was calculated using a base temperature of 10 C (Leong and Ong, 1983).
Samples of 10 randomly selected plants from each subplot were harvested every 10–13 d between R3 and R8 crop growth stages (Boote, 1982). After that, the plants were separated into their components (stem, leaves, pods and seeds) and classified into the main stem (n), cotyledonary branches (n+1 cot), other n+1 branches, n+2 cotyledonary branches (n+2 cot), other n+2 branches, and, eventually, n+3 branches. Finally, plant materials were dried in an oven at 70–80 C for at least 72 h, and dry weights were measured.
With this plant biomass data, crop growth rate (CGR) during the effective pod-filling period was estimated as the quotient between plant biomass increase and the duration of the period; a partitioning coefficient (p) was calculated as the ratio of pod growth rate (PGR) to CGR. From the curves of number and weight of pods, the rate, starting, ending, and duration time were estimated using a bilinear model with plateau (Eq. (1) and (2)) with the nonlinear routine of Table Curve 2D software, Version 5.01 (Jandel TBLCURVE, 2008).
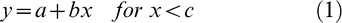
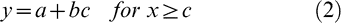
where a is the intercept, b the slope, c the breakpoint of the functions, x thermal time after sowing, and indicates the value of x for which y is at its maximum.
The bilinear model with plateau was plotted for number (a) and weight (b) of pod for each branch type as a function of thermal time after sowing, while the rate of these processes (parameter b) and its duration was calculated as the period between parameter x, when y is zero, and parameter c (Tanaka and Maddonni, 2009). After that, the correlations between CGR, p, IPAR and mean temperature prevailing during the period of pod number and pod weight formation for each branch type were calculated.
Results obtained were subjected to analysis of variance (ANOVA), means were separated using Fisher's protected least significance difference (LSD) test at 95% level of probability, and Pearson's correlation coefficients were computed among some variables using INFOSTAT software (Di Rienzo et al., 2011).
Results and Discussion
During the peanut growing season 2009/2010, temperature and IPAR values recorded were similar to those described by Seiler et al. (1995) for a 30-year period of climatic data.
The total number and weight of pods obtained at harvest (R8 stage), classified according to branch type, are presented in Tables 1 and 2, respectively. Granoleico showed higher number (49 pods/plant) and weight (38.7 g/plant) of pods than Utre (40 pods/plant and 31.4 g/plant) when analyzing the three sowing dates. Both parameters showed decrease as the sowing date was delayed, which was observed for all branch types but with no differences between them.
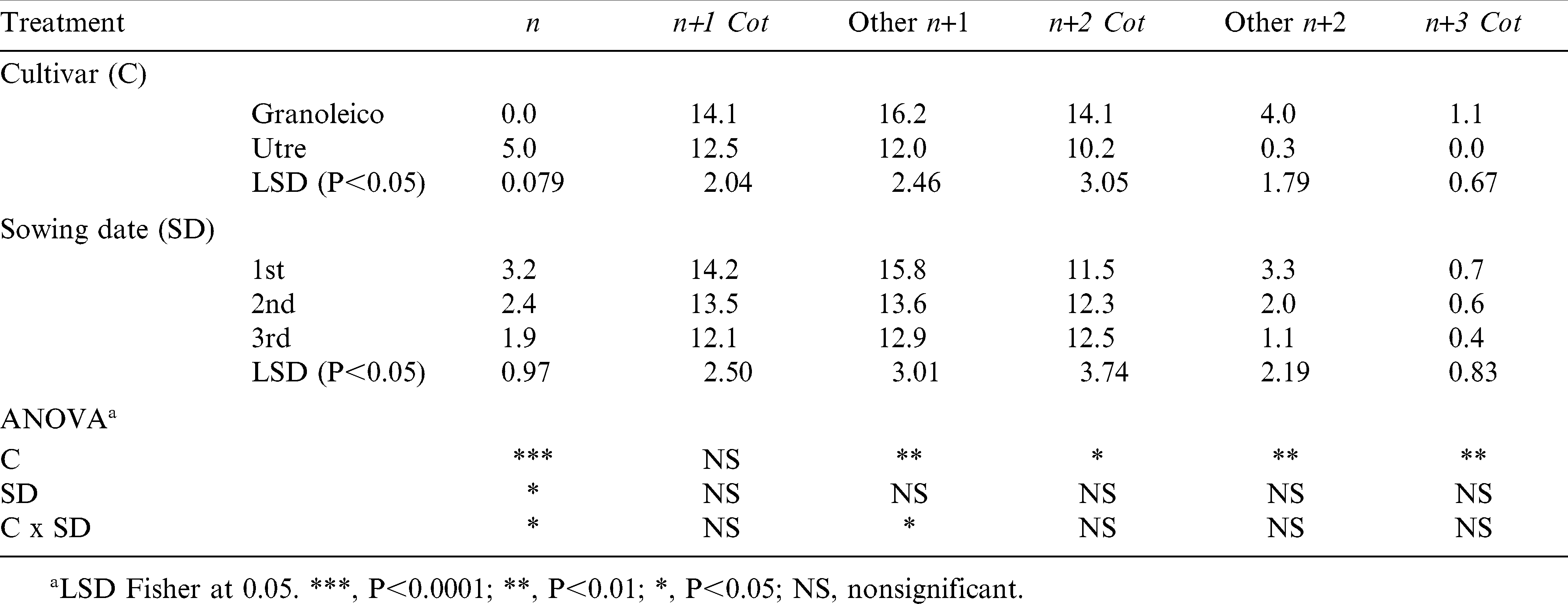
Total pod number per plant in each type of cultivar at R8 phenological stage for all branch types and sowing dates.
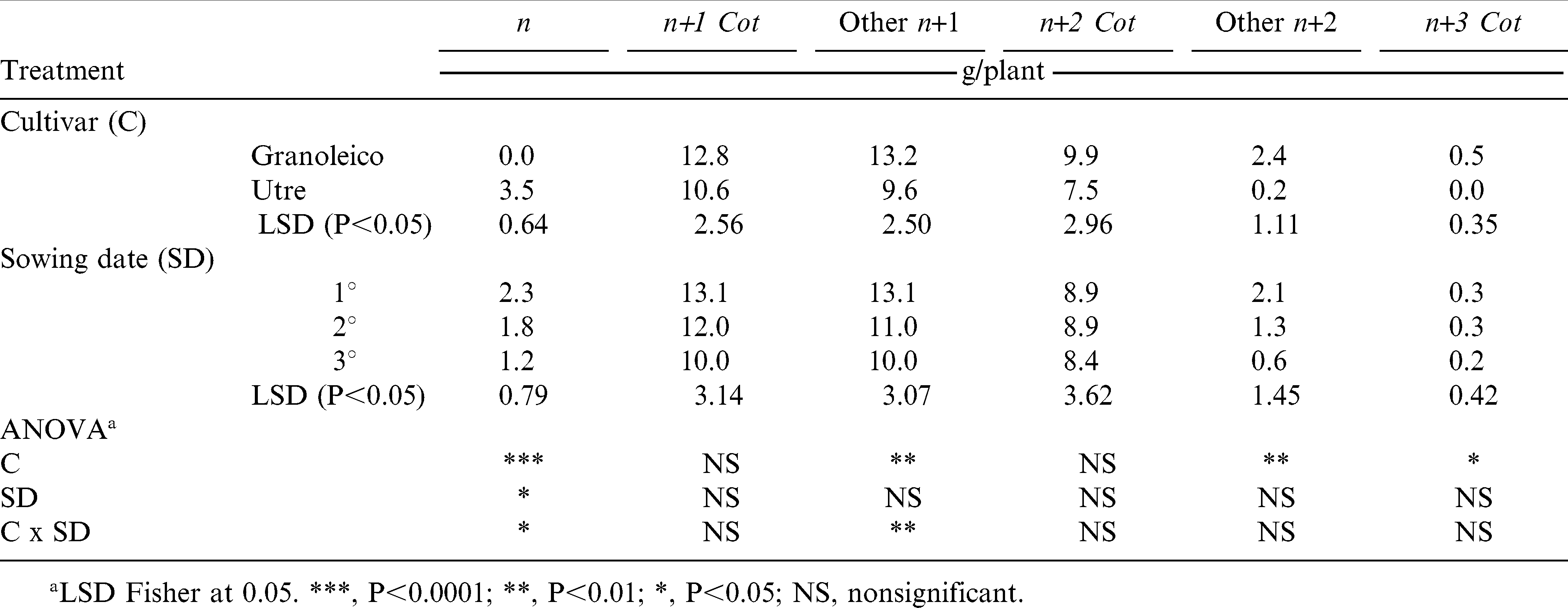
Total pod weight per plant in each type of cultivar at R8 phenological stage for all branch types and sowing date.
Branch type contribution to crop yield varied with the cultivar, but did not vary with the weather conditions in each sowing date. The genotypic regulation on biomass distribution among the different growth habits and the branching patterns of cultivars (Shashidhar et al., 1986; Bell et al., 1993; Giayetto et al., 2007) appeared to be of greater importance.
The number and weight of pods was higher in cotyledonary n+1 of Utre and in other n+1 of Granoleico, in agreement with results obtained by other studies (Choudhari et al., 1985; Shashidhar et al., 1986; Giayetto et al., 2007). Thus, the n+1 branches (both cotyledonary and others) accounted for 61% of the total number of pods per plant in both cultivars, and 65 to 67% of pod weight in Utre and Granoleico, respectively. These values are lower than those reported by other authors: 86 to 91% (Choudhari et al., 1985), 85% (Shashidhar et al., 1986) and 73% (Giayetto et al., 2007). However, the contribution to plant yield of n+2 cot branches reported for this study (24 to 26% of pod weight for Utre and Granoleico, respectively) was greater than in studies cited above. Shashidhar et al. (1986) also noted that spanish genotypes with presence of n+2 cot produced greater pod yields. Thus, the n+1 cot, other n+1 and n+2 cot branches explained an 89 and 93% (Utre and Granoleico, respectively) of pod yield. Utre had an additional 11% contribution to the number and weight of pods from the main stem (n), with only a minor contribution of other n+2 branches (0.5%); whereas, in Granoleico the other n+2 contributed with an additional 6% and n+3 cot with 1%, thus highlighting the different branching patterns as described by Stalker (1997).
The critical period for the pod number determination in the main categories of branches (n+1 cot, other n+1 and n+2 cot) began at R2 and R3 and lasted until stages R5 and R6 (pod filling) (Table 3). The duration of this period was 600 growing degree d (Cd) for Granoleico and 436 Cd for Utre. The differences between cultivars were due to the growing season duration (longer in the runner-type) (Giambastiani, 2000). The earlier onset and longer duration occurred in cotyledonary n+1 branches of Utre and in the other n+1 of Granoleico. In contrast, the highest rate of pod set occurred in n+1 cot, other n+1, and n+2 cot branches of the two cultivars (Figure 1).
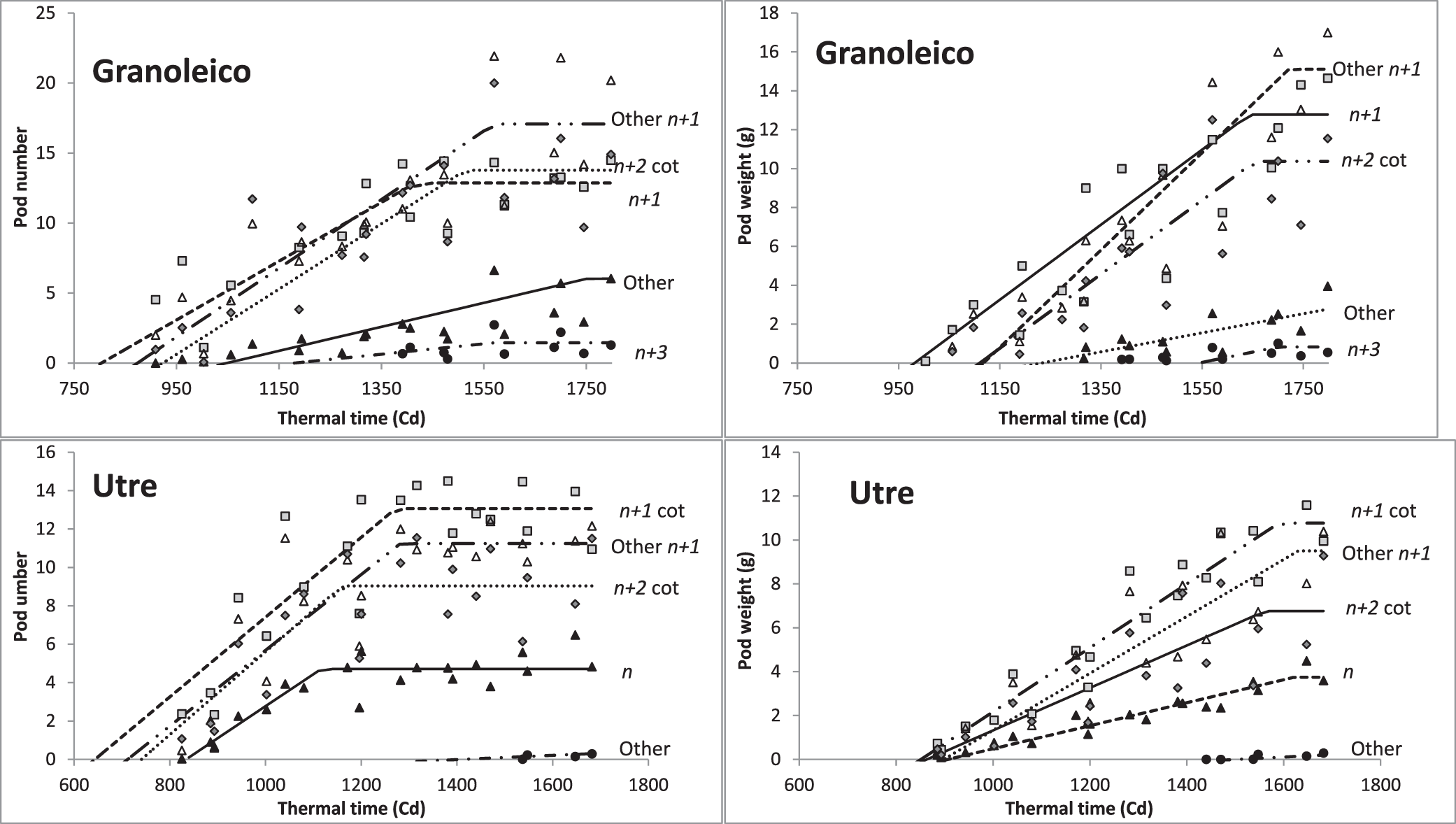
Bilinear relations between number and weight of pods (g) and thermal time (Cd) after sowing (Tb: 10 C) for Granoleico and Utre cultivars.
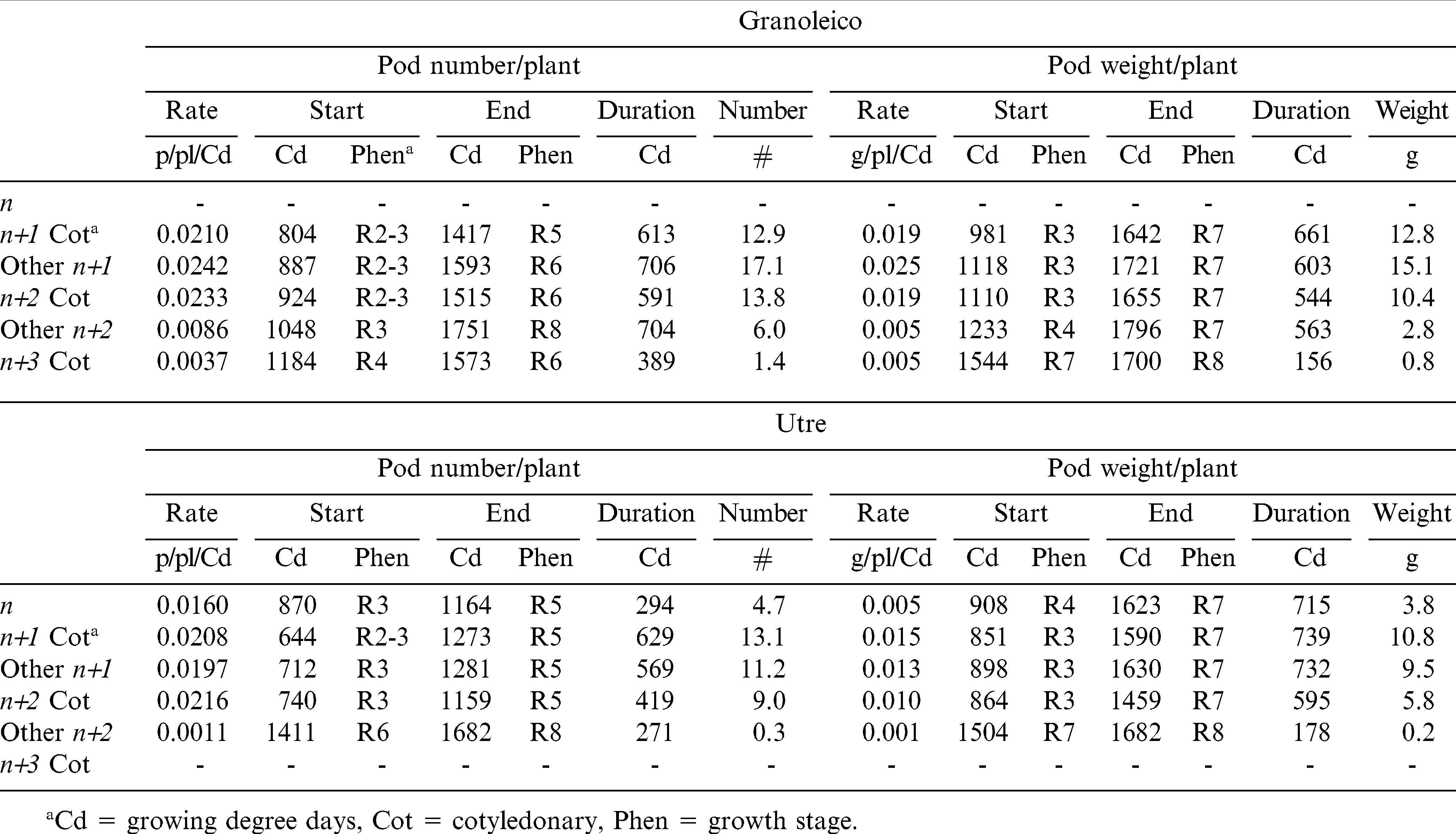
Starting, ending, duration in Cda, rate of appearance (pods/Cd), and pod growth (g/plant/Cd) during the linear growth by branch type.
In Granoleico, critical periods of pod weight determination followed a pattern similar to that described for pod number, with three groups: cotyledonary n+1 branches, other n+1 and n+2 cot, and, finally, a group formed by the other n+2, and n+3 cot branches. Differences between groups were due to the PGR and the duration of the stages. In Utre, the beginning and end of the determination period of pod weight were similar in the main branch types (n+1 cot, other n+1, and n+2 cot), and differences founded in the final pod weights were mainly due to PGR (Figure 1). The genotypic differences observed should be due to the degree of indeterminacy characteristic of each cultivar.
Tables 4 and 5 present average weather conditions for the three sowing dates, the crop growth rate (CGR), and the partitioning coefficient (p) for the critical periods of number and weight of pods in each branch type. The average daily intercepted radiation (IPAR) by the crop canopy showed no significant differences, except during the fruiting period of Utre other n+2 branches. A similar situation was observed for the mean temperature during pod weight determination and for CGR in both periods. This was due to the fact that other n+2 branch type fructified late in the crop growing season. Air temperature and IPAR showed no statistically significant differences among branch types for the Granoleico cultivar. The partitioning coefficient of pods was the only variable that differed significantly between branch categories in both cultivars for pod number and weight formation periods. Thus, the higher value of p during number and weight determination periods corresponded to cotyledonary branches in Utre and to other n+1 branches in Granoleico. On other hand, the partitioning coefficient (p) of each branch (Tables 4 and 5) was related to their respective final data of pod number and weight (Tables 1 and 2).
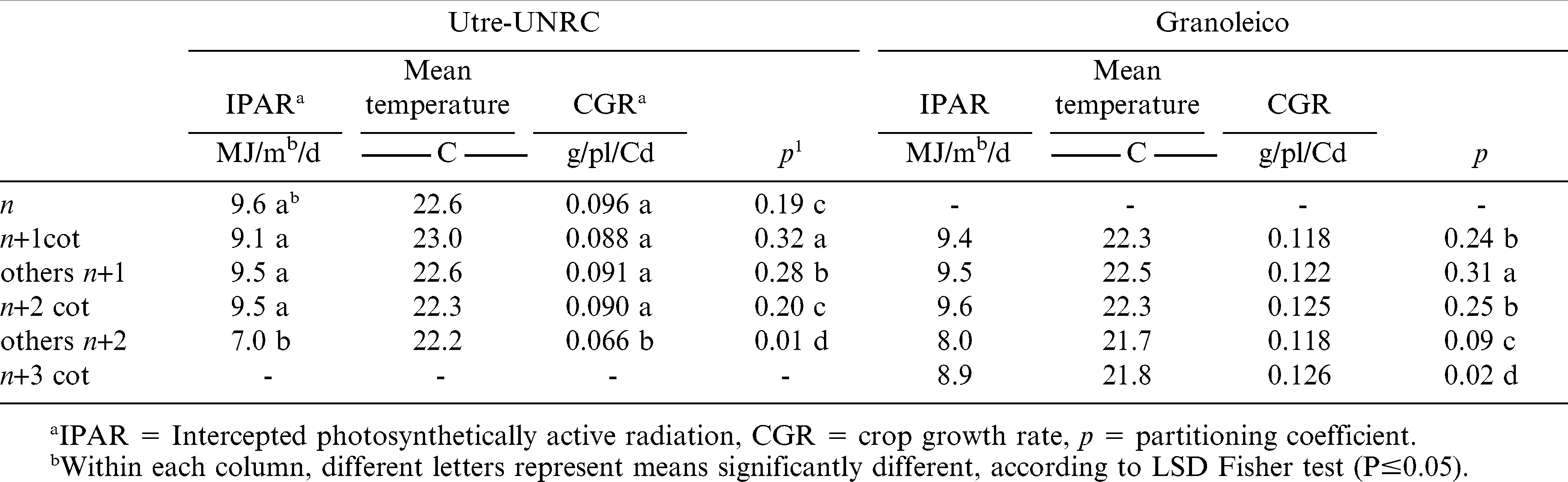
Intercepted photosynthetically active radiation, mean temperature, crop growth rate and partitioning coefficient for each branch type during the period of pod number determination.
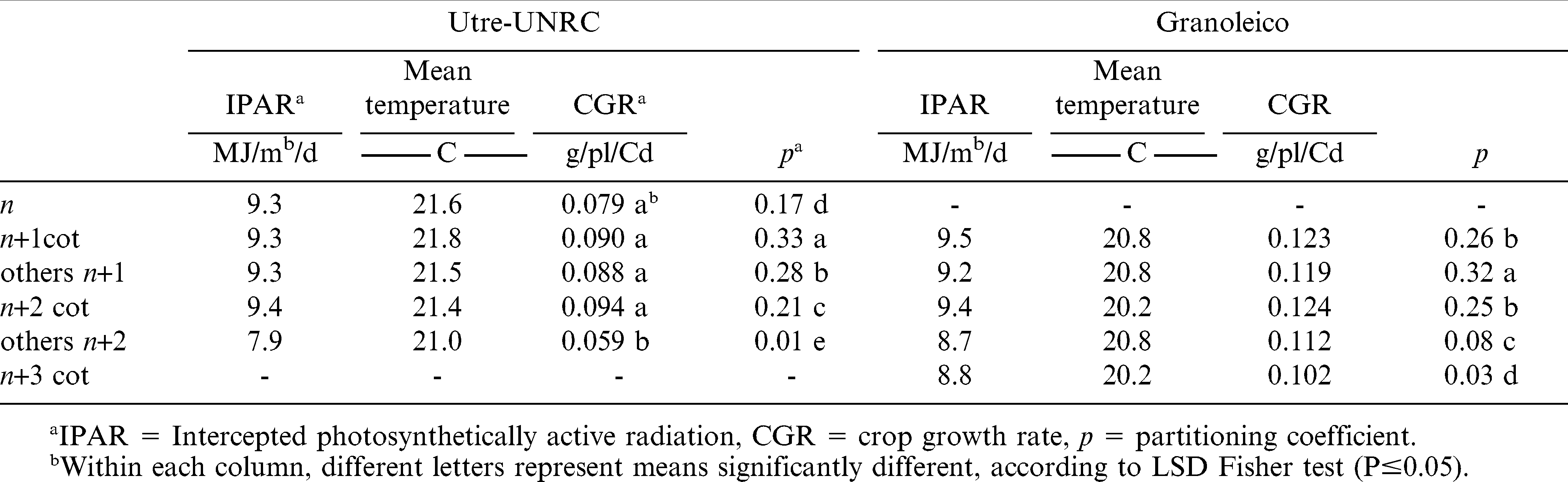
Intercepted photosynthetically active radiation, mean temperature, crop growth rate and partitioning coefficient for each branch type during the period of pod weight determination.
The correlations between pod number and pod weight and partitioning coefficient (p) of branches for both cultivars during their respective determination critical period were highly significant (Table 6). However, we found no significant correlations with the mean air temperature and IPAR evaluated during the periods of number and weight of pod determination. This might be because the critical periods of yield component determination for each branch type were not very different from one another (similar occurrence and duration). Thus, the yield components determination critical periods were exposed to similar weather conditions. This result contrasted the results found by Bell and Wright (1998).
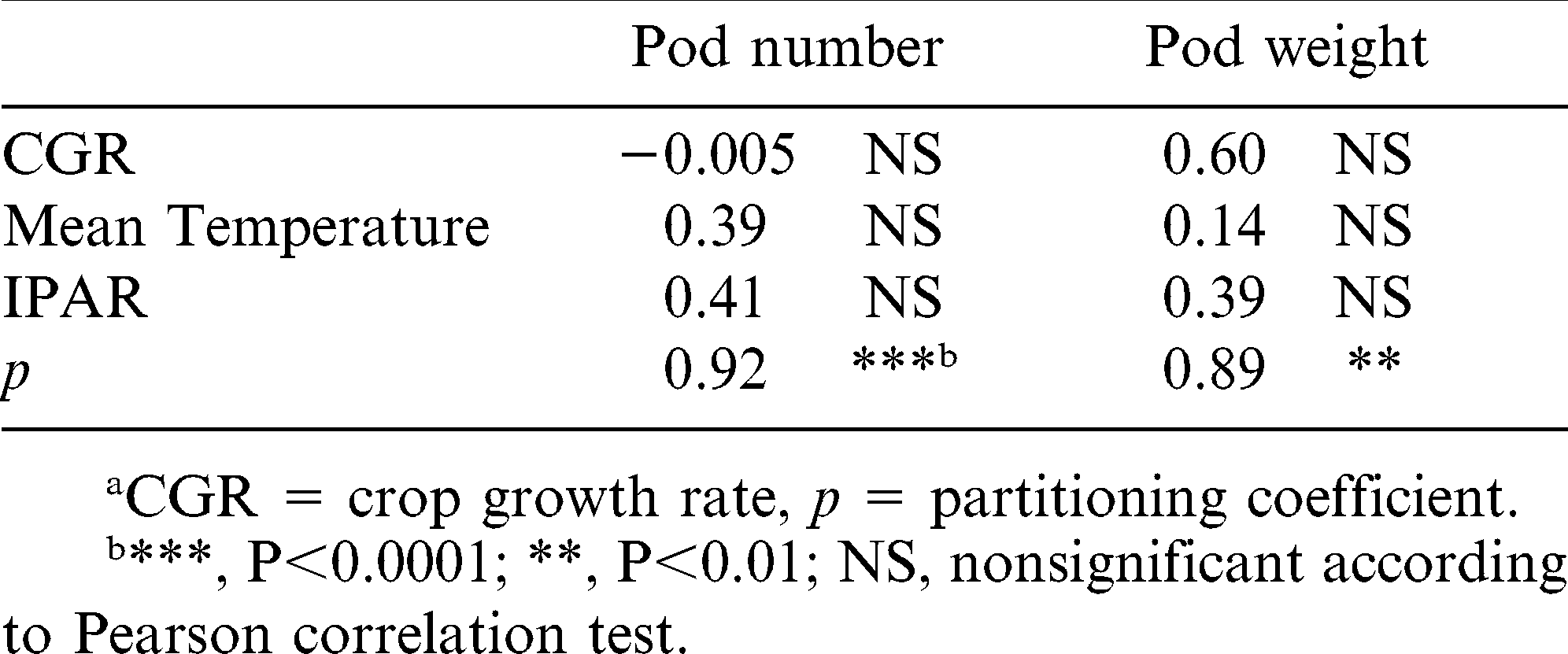
Correlation coefficients among number and weight of pods and weather variables, crop growth ratea, and partitioning coefficienta during the periods of number and weight of pods determination.
The correlation between pod number and CGR was negative due to at the beginning time of pod number determination on the branches with greater contribution to pod yield the CGR was low. However, when the determination of pod number began on branches of minor contribution, the CGR was higher. Conversely, during the period of pod weight determination there was a positive correlation with CGR. These results are consistent with those noted by Bell et al. (1993) for the total plant.
It is likely that the variation found was due to the sink strength (number and weight of pods) which depends, among other things, of the pod position in the plant, being this attribute determined by both the branch type as by the position of reproductive nodes on it (Bell et al., 1993). Thus, the branches that develop earlier, located in proximity to the ground, will be the first to start defining the number and weight of pods (Choudhari et al., 1985; Shashidhar et al. 1986) and with them the sink strength. Therefore, these branches will show a high partitioning coefficient thus achieving higher yields than those with a later ontogeny.
Conclusions
These data indicate the differences in the relative contribution to crop yield provided by each branch type are due to the critical period of determination of number and weight of pods. Critical periods differ between genotypes, these periods being dependent on growth habit and branching pattern; branches with greater contribution to crop yield have an earlier onset, higher rate of appearance and pod growth, and longer duration of those critical stages. However, there was no difference in the amount of IPAR and average temperature due to temporal overlap of these stages in the main branch types (n+1 cot, other n+1, and n+2 cot). Instead, the partitioning coefficient was differential among branch types, and would be defined by the sink strength (number and weight of pods), which in turn, are higher in the branches that first define the yield components, giving these branches a comparative advantage with respect to the later ontogenic development branches.
Acknowledgements
Authors wish to thank Silvia Beck for the help with manuscript translation. Federico D. Morla has a graduate scholarship from the National Council for Research of Argentina (CONICET) and this work is part of his doctoral thesis.
Literature Cited
Bell M.J and Wright G.C 1998 Groundnut growth and development in contrasting environment. 2. Heat unit accumulation and Photo-Thermal effects on Harvest Index Expl. Agric. 34 ( 1 ): 113 – 124 .
Bell M.J Wright G.D and Harch G 1993 Environmental and Agronomic Effects of four Peanut Cultivars in a Sub-Tropical Environment. II. Dry Matter Partitioning Expl. Agric. 29 : 491 – 501 .
Boote K.J 1982 Growth Stages of Peanut (Arachis hypogaea L.) Peanut Sci. 9 ( 1 ): 35 – 40 .
Choudhari S.D Udaykumar M and Sastry K.S.K 1985 Physiology of bunch groundnuts (Arachis hypogaea L.) The Journal of Agricultural Science. 104 ( 02 ): 309 – 315 .
Di Rienzo J.A Casanoves F Balzarini M.G Gonzalez L Tablada M and Robledo y C.W 2011 InfoStat versión 2011 Grupo InfoStat, FCA, Universidad Nacional de Córdoba , Argentina .
Giambastiani G Rendimiento de dos cultivares de maní de distinto ciclo en diferentes épocas de siembra 2000 In XV Jornada Nacional de maní. Gral. Cabrera, Córdoba .
Giayetto O Fernandez E.M Bessone A and Cerioni Y.G.A 2007 Patrones de ramificación, crecimiento de frutos y calidad de genotipos de maní In XXII Jornada Nacional de Maní, 1er Simposio de Maní en el MERCOSUR General Cabrera , Córdoba .
Jandel TBLCURVE 2008 TableCurve Curve fitting software. Jandel Scientific. Corte Madera, CA .
Leong S.K and Ong C.K 1983 The influence of temperature and soil water deficit on the development and morphology of peanuts (Arachis hypogaea L.) J. Exp. Bot. 34 : 1551 – 1561 .
Rehman A.U Wells R and Isleib T.G 2001 Reproductive Allocation on Branches of Virginia-Type Peanut Cultivars Bred for Yield in North Carolina Crop Sci. 41 : 72 – 77 .
Seiler R.A Fabricius R.A Rotondo V.H and Vinocur M.G 1995 Agroclimatología de Río Cuarto – 1974 / 1993, U.N.R.C. Vol. I. 68 .
Shashidhar V.R MalathiChari, Prasad T.G and Udaya Kumar M 1986 A Physiological Analysis of the Branching Pattern in Sequential Types of Groundnut in Relation to the Fruiting Nodes and the Total Mature Pods Produced Ann Bot. 58 ( 6 ): 801 – 807 .
Sholar J.R Mozingo R.W and Beasley J.P 1995 Peanut cultural practices . pp. 354 – 382 In Pattee H.E and Stalker H.T (Eds.) Advances in Peanut Science Stillwater, OK, USA .
Stalker H.T 1997 Peanut (Arachis hypogaea L.) Field Crops Res. 53 : 205 – 217 .
Tanaka W and Maddonni G.A 2009 Maize Kernel Oil and Episodes of Shading during the Grain-Filling Period Crop Sci. 49 ( 6 ): 2187 – 2197 .
Notes
- Departamento Producción Vegetal, Facultad de Agronomía y Veterinaria, Universidad Nacional de Río Cuarto. Ruta Nacional 36, Km 601. 5800 Río Cuarto, Córdoba Argentina * Corresponding autor, email: ogiayetto@ayv.unrc.edu.ar
Author Affiliations