

Introduction
Peanut (Arachis hypogaea L.) is an economically valuable crop that is cultivated extensively worldwide. A substantial market exists for in-shell peanuts, with shell color being one of the factors affecting price: a bright-yellow color is favored by consumers (the premium for bright shells is 30% higher). In Israel, peanuts were grown mainly in sandy soils; when the cultivating area was expanded to land that contains loess soils, there was a loss of the shell's marketable yellowish color to a less desirable yellow-brown tint. As peanut pods develop underground, the effect of soil type on shell color was studied.
The peanut plant produces its flowers aerially. Upon fertilization of the ovule in the flower, a specialized organ, the gynophore (peg), begins to form and grow downward, pushing the ovary into the soil, where pod maturation occurs. While exposed to air, the embryos are in an arrested state of development; it is only after the ovule region of the gynophore is pushed below the soil surface and positioned parallel to it that the pod begins to grow.
The peanut shell originates from the ovary wall. The anatomy of the peanut pod, including shell development, has been described in detail by Periasamy and Sampoornam (1984) (see also Figure 2). The mature shell consists mainly of inner papery white tissue originated from the innermost layer of the ovary, a fibrous tissue composed of branched vascular bundles interconnected with lignified tissue, and an external tissue composed of a few layers of thick-walled corky cells (Periasamy and Sampoornam, 1984; Xu et al. 2000).
It is highly likely that chemical composition of peanut shells varies with peanut variety and shelling conditions (Colwell et al. 1945); however, no direct evidence has been found for the effects of a physical interaction between shell material and the surrounding soil. The pod shell contains mainly cellulose, hemicellulose and lignin, as well as crude protein, ash and fat (Pickett, 1950; Wartelle and Marshall, 2000). The developing shell also contains sucrose, starch and water; however, towards maturation, hemicellulose content increases while the percentage of all non-fibrous components decreases (Pickett, 1950). The fibrous component of peanut shells has the capacity to adsorb toxic metals (Henderson et al. 1977); heat or chemical treatments are currently being used to improve their ability to uptake metal ions and organic compounds from industrial waste (Wartelle and Marshall, 2000; Chamarthy et al. 2001; Wilson et al. 2006). This characteristic of the shell material may play a role in adsorbing a soil component(s) that produce the final pod color.
The aim of the present study was to establish the relationship between shell color and soil type, and to define the pod developmental stage at which shell color is determined. Results suggested that a water-soluble soil factor(s) and a pod surface component(s) are involved in determining the shell's final color. Reducing soil wetness at the end of the growing period (3–4 weeks prior to harvest), when the pod is maturing, may contribute to maintenance of the commercially desirable bright-yellowish color of the shells.
Materials and Methods
Plant Material and Growth Conditions
Cultivar Hanoch of the Virginia-type peanut was used in the present study. Field experiments were performed in the western Negev area of Israel (N31°26.1768, E34°29.9064) during the growing seasons (Apr–Sep) of 2007 and 2008. Plants were sown in three rows in a 70-cm wide bed. Cultural practices (i.e., fertilization, irrigation, peanut drying, etc.) were followed as recommended for this region.
Soils for experiments were chosen based on initial shell (pericarp) color, which was obtained in preliminary observations (Table 1). In greenhouse experiments, seeds were planted in 50-L pots filled with sand1.6 or loess2 soils. At sowing, seeds were inoculated with a mixture of local peanut-specific Bradyrhizobium strains (280A, 2209A, R283A1) at a final concentration of 2.9 × 107 cells/ml (2 ml/seed). Plants were irrigated with tap water which was supplemented twice weekly with N-free nutrient modified Johnson solution (Johnson et al. 1957). Prior to collecting measurements peanuts were baked in an oven at 70 C for 8–10 days to ensure complete dryness.
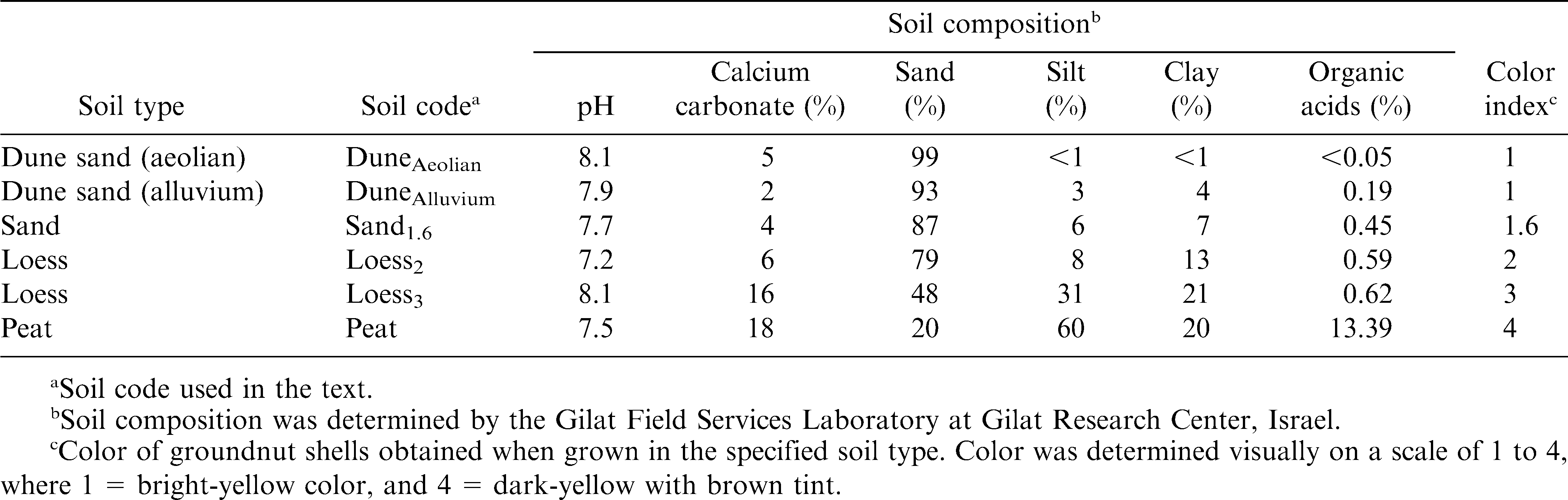
List of soils used.
Shell Color Determination
Peanut growers determine shell color visually, on a scale of 1 to 4, where 1 = bright yellow and 4 = dark yellow with brown tint. In the present work, the color was determined using a portable spectrophotometer spectro-guide CC-6800 (BYK-Gardner, Columbia, MD) and based on the L*a*b* model that presents color in a three-dimensional space. The three basic coordinates represent the lightness of the color (L*, L* = 0 yields black and L* = 100 indicates white), its position between red/magenta and green (a*, negative values indicate green while positive values indicate magenta), and its position between yellow and blue (b*, negative values indicate blue and positive values indicate yellow). The color value for each peanut pod was given by an average of five readings at different locations on the shell surface; samples consisted of 12 to 25 pods. Treatments were performed in three or more replicates as described further on. Data were analyzed for statistical significance among means (P > 0.05) by Tukey HSD test (JMP software, http://www.jmp.com).
A comparison of visual color evaluation and spectro-guide readings indicated a strong correlation between the methods: for brightness (L* values), y = −2.3971x + 74.657, r2 = 0.9862; for red tint (a* values), y = 1.2037x + 2.315, r2 = 0.9646; for yellow tint (b* values), y = −0.7024x + 32.578, r2 = 0.9151). These correlations indicate strong support for the spectrometric method in determining shell color.
The Effect of Soil Extracts on Shell Color
Bright-yellow mature pods (color index = 1) were incubated overnight at 4 C in autoclaved-sterile saturated test-soil solutions (50% w/v in water), in filtered extract of the same test-soil solutions, or in water. Test soils were duneAeolian, loess3 and peat. Following the incubation, pods were washed lightly with tap water, dried and their color was determined. Experiments were performed in triplicates, 12 pods per replicate.
Field and Greenhouse Experiments
Experiment 1: The Effect of Soil Type on Shell Color
Plants were grown in sand1.6 soil while the soil between plant lines in the plot was replaced with test soils, where pods were allowed to develop. Each replacement area was 30 cm wide (space between plant lines), 30 cm long and 20 cm deep, and was lined at its perimeter with plastic sheets (see Figure 1A). Soil replacement was replicated nine times for each soil type (Table 1) in a complete randomized block design. A similar experiment was set up in a greenhouse where plants were grown in pots filled with sand1.6 while pods were directed to develop in neighboring pots filled with the test soils (see Figure 1C,D). At the end of the growing season, mature peanut pods were collected for shell color determination.

The effect of soil type on shell color. Peanut plants were sown in sandy soil, but their pods were directed to develop in various soil types (Field experiment 1). (A) Field experiment, in which local soil between the rows was replaced with the test soils; (B) 60 days later foliage covered the experimental plots. (C) Greenhouse experiment, in which plants were sown in a central pot and (D) gynophores were directed to pots containing the test soils. (E) Mature pods collected from the above experiments, demonstrating the effect of soil type on shell color. 1, duneAlluvium; 2, duneAeolian; 3, sand1.6; 4, loess2; 5, loess3; 6, peat.
Experiment 2: Soil Wetness and Shell Color
Plants were grown in a loess3 peanut field, in two rows (70 cm apart) in a raised bed (width 1.93 m), 7 plants per 1 m row. Up to 100 days post-sowing (DPS) (seed-filling stage), the experimental plots were irrigated by sprinklers every 6–7 days with 35 mm of water; total amount of water per plot for that growth period was 480 mm. From that time on, three irrigation protocols were applied according to water potential values measured by tensiometers set at a depth of 25 cm below the soil surface: (a) conventional field practice — 35 mm water given every 6–7 days when soil water potential reached 50 centibar (added amount of water 285 mm, total amount of water for the entire growing season 765 mm), (b) 35 mm water given every 10 days when soil water potential reached 80 centibar (added amount of water 215 mm, total amount of water for the entire growing season 695 mm) (“dry” treatment), and (c) 50 mm water given every 10 days when soil water potential reached 50 centibar (added amount of water 295 mm, total amount of water for the entire growing season 775 mm) (“wet” treatment). Each plot consisted of three beds at a length of 200 m. Treatments were replicated three times in a complete randomized block design. At the end of the growing season and the test period, mature peanut pods were collected for shell color determination.
Experiment 3: Control of Soil Wetness in the Pod-Development Zone
Plants were grown in a loess3 peanut field, in two rows (70 cm apart) in a raised bed (1.93 m width), 7 plants per 1 m row, and were watered by subsurface drip system, one extension per bed. Dripper lines were buried at 5 cm (designated ‘drip5’) or at 25 cm (‘drip25’) below the soil surface. ‘Drip25’ supplied water directly to the root zone, while ‘drip5’ watering flowed to the roots via the pod-development zone. In another treatment, both ‘drip5’ and ‘drip25’ extensions were buried in the plot; plants were watered by ‘drip5 up to 100 DPS and then the system was changed to ‘drip25’ irrigation. Conventional sprinkler irrigation was used as a control. Tensiometers were set at a depth of 25 cm under the soil surface. Watering was operated (25–30 mm) when soil water tension reached 40 centibar. Drip treatments received a total 671 mm of water while sprinkler irrigation controls got 761 mm. Each plot consisted of three beds at a length of 200 m. Treatments were replicated four times in a complete randomized block design. At the end of the growing season, mature peanut pods were collected for shell color determination.
Experiment 4: Identifying the Pod Developmental Stage Most Sensitive to Soil Wetness and Loss of Shell Color
Plants were grown in a loess3 peanut field, in two rows (70 cm apart) in a raised bed (1.93 m width), 7 plants per 1 m row. The experimental area included six raised beds with one extension of a 25-cm deep subsurface dripper line per bed. The experimental area was divided lengthwise into five plots, 24 m each, and valves were installed on the dripper lines every 24 m, accordingly. First, plants were watered by sprinklers; however at various times post-sowing (85, 100, 120 and 140 days), the sprinklers in sequential plots were stopped and the drip system was operated. The last plot, used as a control, was irrigated by sprinklers throughout the entire growing season and represented the practical recommendations for this region. Total amount of water was about 761 mm in all treatments and control. At the end of the growing season, mature peanut pods were collected for shell color determination. Due to technical difficulties, the experimental plots could not be arranged in a randomized design and therefore samples were collected at six locations in the beds from each treatment (in-line replicate design).
Anatomy of Shell Development
Peanut pods were collected at various developmental stages. Tissue samples (blocks of 4 × 3 × 3 mm) were fixed in FAA (50% ethanol, 5% acetic acid and 3.7% formaldehyde, v/v), dehydrated in an ethanol/Histoclear (Finkelman Chemicals, Petach-Tikva, Israel) series and embedded in paraplast (Paraplast Plus, McCormick Scientific, St. Louis, MO) according to standard methods (Ruzin, 1999). Tissue sections (15–20 µm) were stained with Safranin-O/Fast green (Sigma Chemicals, Rehovot, Israel) for examination of tissue morphology (Johansen, 1940). Sections were observed under a light microscope (Leica DMLB, Wetzlar, Germany) and images were displayed on a monitor through a CCD camera (Leica DC2000) using the Leica IM1000 program.
Results
The Effect of Soil Type on Shell Color
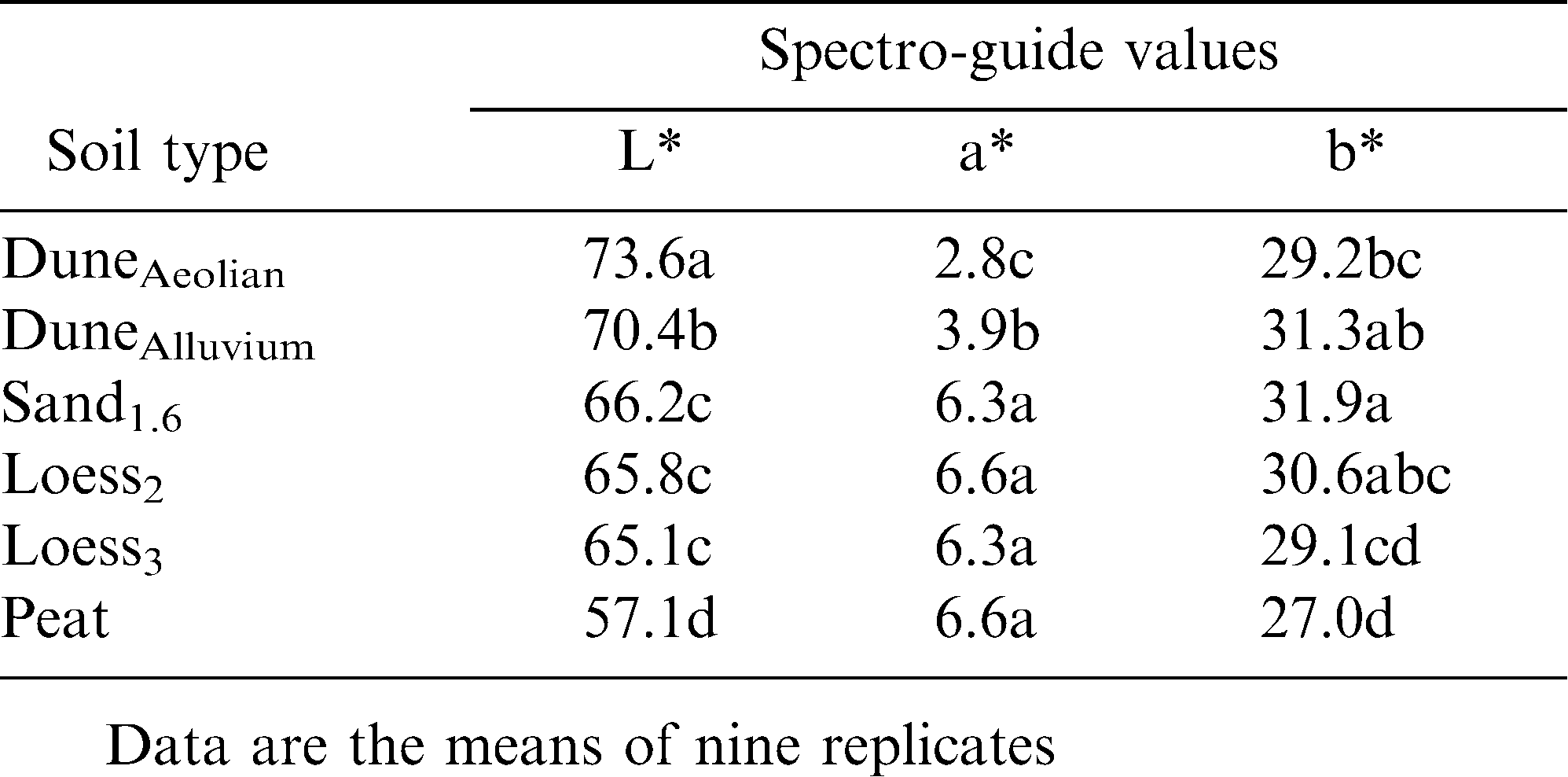
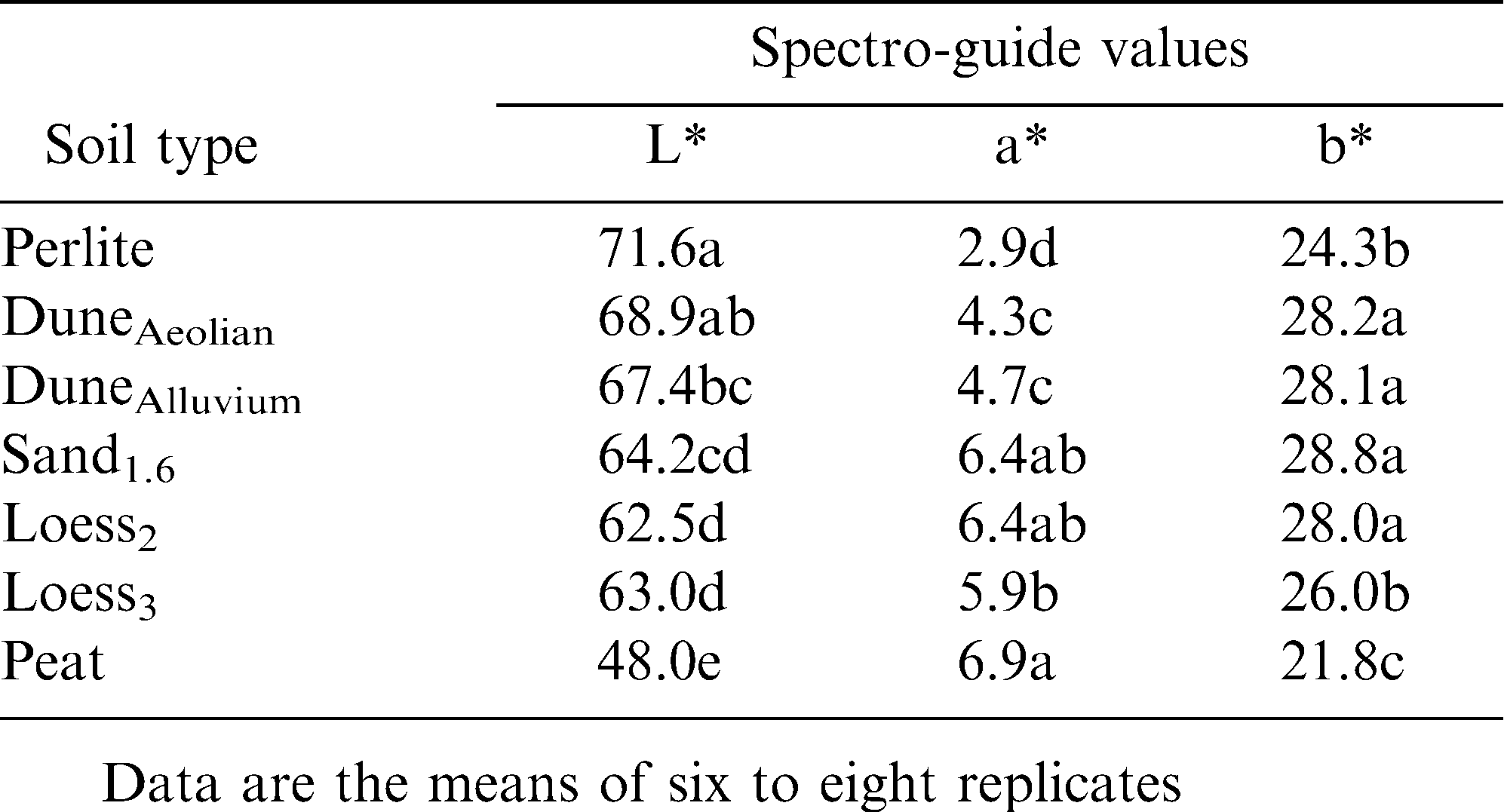
To evaluate the effect of soil on final shell color, experiments were conducted in which the pods of an individual plant were allowed to develop in various test soils (Field experiment 1). Two experiments were conducted in parallel: a field trial in which the soil between rows of plants was replaced by the test soils (Figure 1A,B), and a greenhouse experiment in which small pots with test soils were placed around the plant (Figure 1C,D). Pods were collected at the end of the growing season and shell color was evaluated visually (Figure 1E), as well as with the spectro-guide device (Tables 2, 3). Both experiments indicated a strong association between soil type and shell color, with sandy soils resulting in brighter shells and pods developed in peat exhibiting a darker tint (Tables 2, 3 and Figure 1E). Pods that developed in loess soils exhibited intermediate shell brightness. It is worth noting that compared to the dark tint (color index = 4), bright yellow (color index = 1) resulted from a reduced level of red component (a*) and an increased level of yellow component (b*) in the color (Tables 2, 3 and the following data).
The association between soil type and shell color may be related to the content of organic acids in the soils (Table 1). Accordingly, pods developed in perlite were significantly bright, almost white, with lowest values for red and yellow color components (Table 3), further indicating the contribution of soil composition to shell color.
Shell color developed in various soil types — Field experiment 1.
Shell color developed in various soil types — greenhouse experiment.
The Effect of Soil Extracts on Shell Color
To test whether soil components adhere to the pod surface, thereby affecting its color, mature peanut pods with a shell color index of 1 were incubated in suspensions of saturated test soils (50% w/v in water). Pods incubated in the peat suspension became significantly darker than non-treated pods, while incubation of the pods in a suspension of duneAeolian gave the brightest shell tint (Table 4). The darker pigment/tint of the former may have been due to the adherence of peat component(s) to the pod surface. Interestingly, the yellowish component of the original shell color was reduced following incubation with each of the soil suspensions (Table 4), implying that water-soluble material is washed off the shell surface. Hence, the experiment was repeated with filtrates of the abovementioned soil suspensions and with water. Shell color from all treatments became brighter, with reduced red and yellow color components (Table 5), supporting the hypothesis of washout of (yellow) material from the surface of the shell.
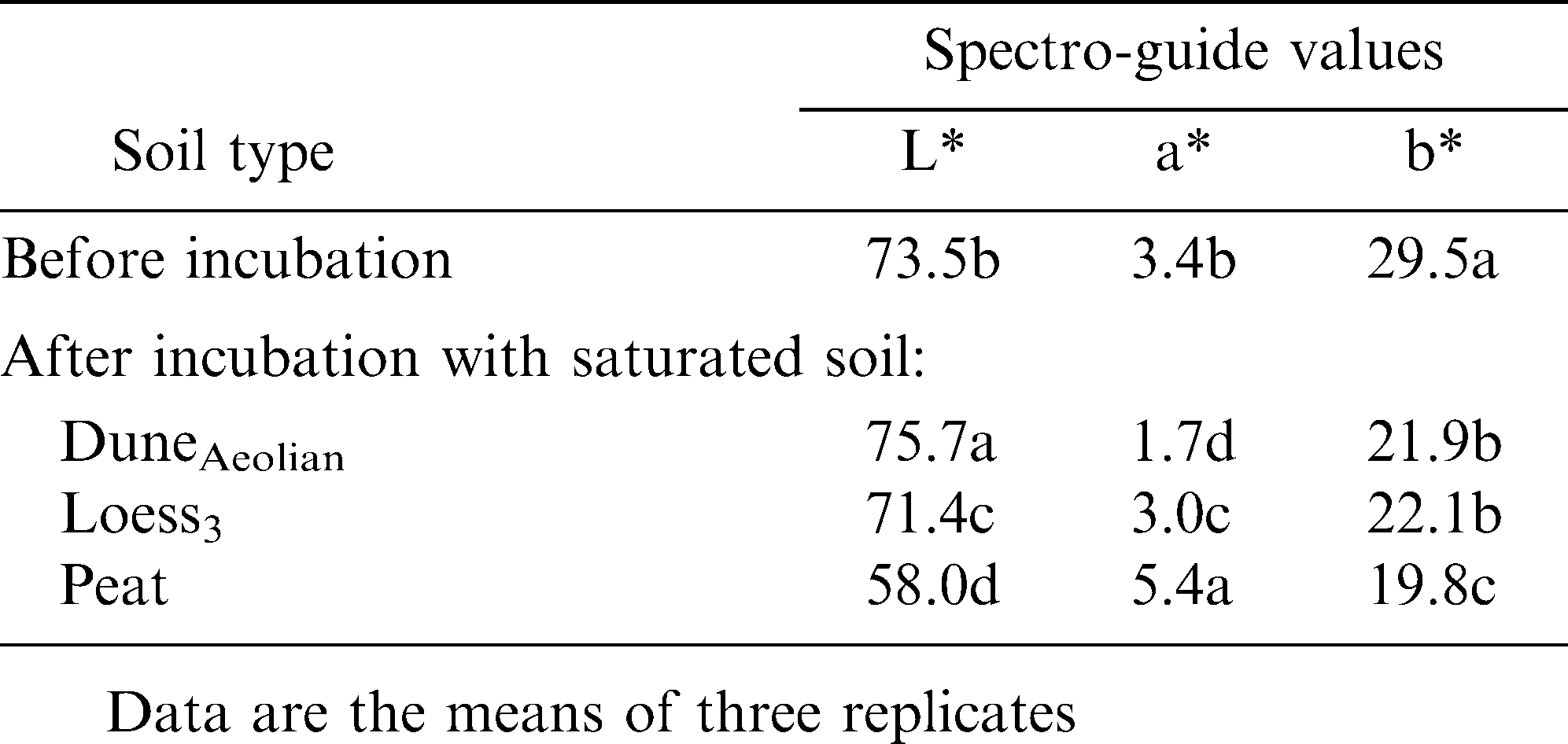
Shell color after incubation with saturated soil suspension.
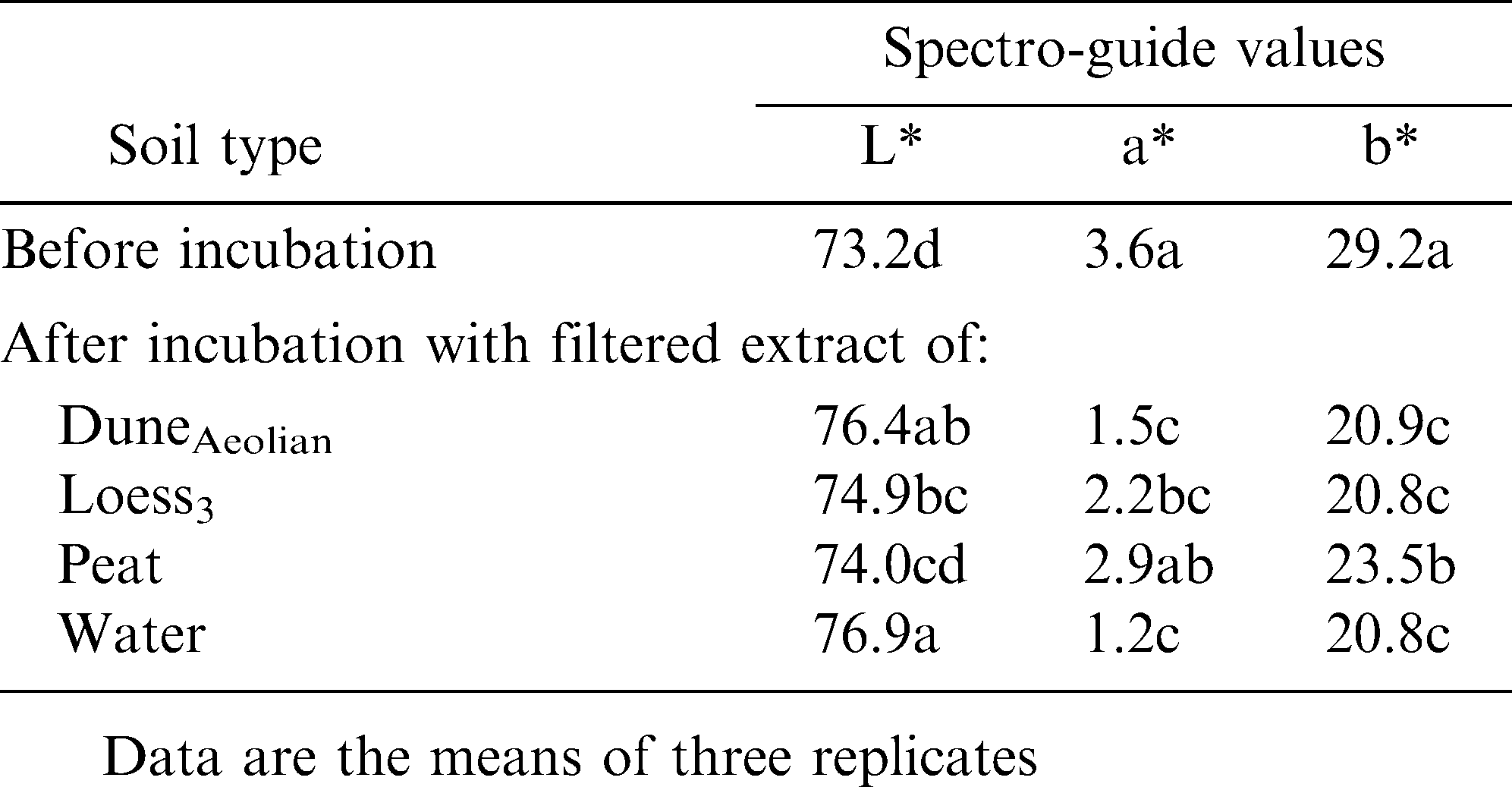
Shell color after incubation with filtrate of soil extract or water.
Soil Wetness and Shell Color
To study the involvement of soil wetness in determining final shell color, a field experiment was conducted in which the soil was kept wet (“wet” treatment), watered with reduced amount of water (“dry” treatment), or irrigated by the conventional growth protocol (Field experiment 2). Shell color from the “dry” treatment was significantly brighter than that from the “wet” treatment; the conventional irrigation protocol resulted in intermediate values (Table 6). These results supported the hypothesis that high wetness of the soil negatively affects shell color. In addition to more desirable shell color, all other pod quality parameters were superior in the “dry” treatment compared to the conventional and “wet” treatments, including total yield, seed size and extent of pathogen infection (O. Buchshtab, unpublished data).

Shell color developed following three sprinkler-irrigation protocols — Field experiment 2.
Control of Soil Wetness in the Pod-Development Zone
Soil wetness in the pod-development zone may be regulated by watering the plant with a drip-irrigation system. The effects of two types of drip systems on shell color were compared: ‘drip25’, in which the dripper line was located in the root zone, and ‘drip5’, in which the dripper line was located just above the pod-development zone. Both treatments were compared to conventional sprinkler irrigation (Field experiment 3). Pods with dark and undesirable tint were obtained when the irrigation water was applied from above the pod-developmental zone — ‘drip5’ and with sprinkler irrigation (Table 7). The brightest shell tint was obtained when plants were continuously irrigated with the ‘drip25’ system or irrigated with ‘drip5’ up to 100 DPS and then with ‘drip25’ until harvest (Table 7). The latter result implied that soil wetness affects shell tint from 100 DPS until harvest.
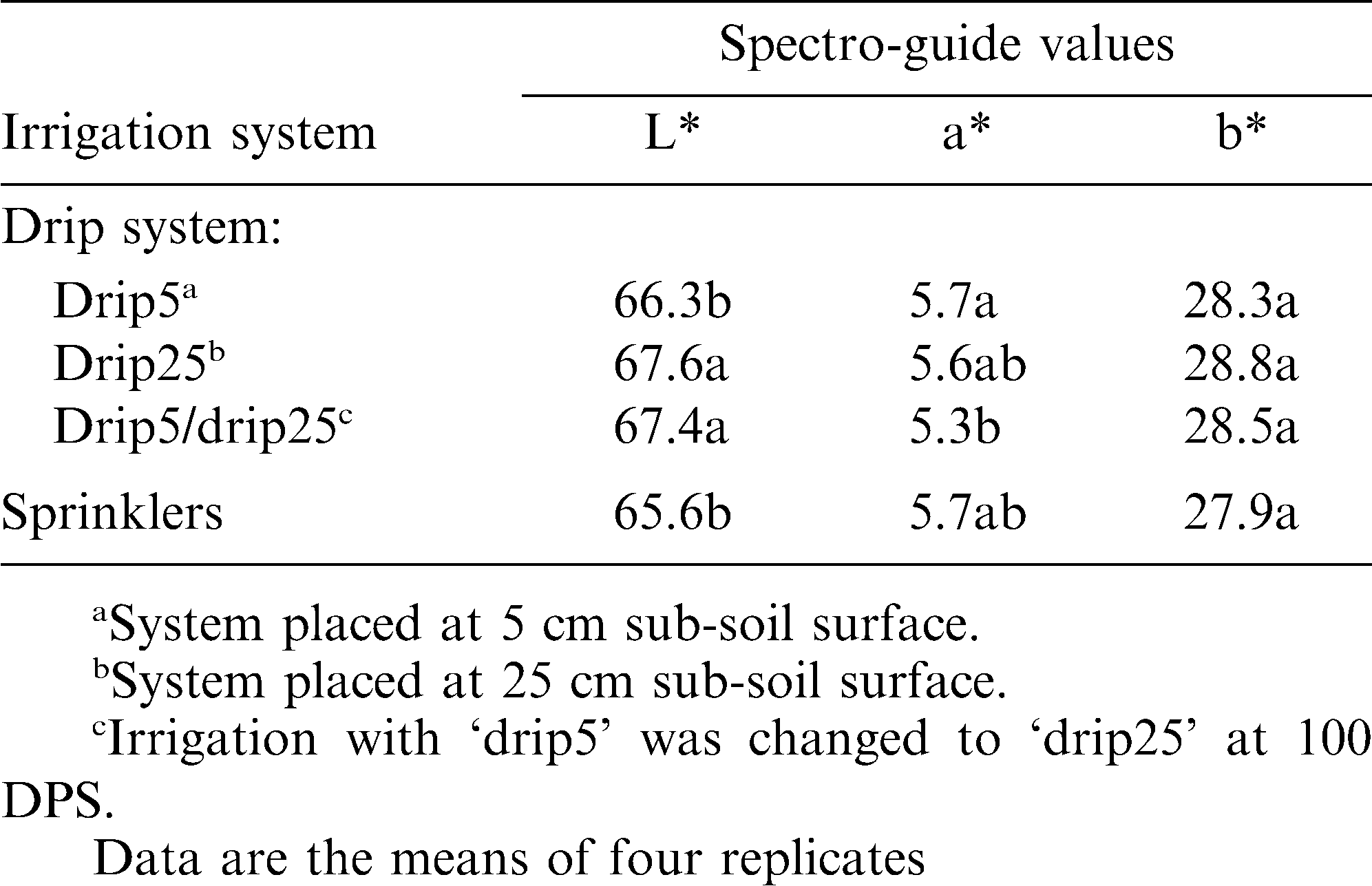
Shell color developed following irrigation with drip systems compared to sprinklers — Field experiment 3.
Identifying the Pod Developmental Stage Most Sensitive to Soil Wetness and Loss of Shell Color
To further determine the developmental stage at which the pod is most susceptible to loss of shell tint, the wetness of the soil surrounding the developing pods was regulated by shifting between sprinkler irrigation and the ‘drip25’ system at several time points after sowing (Field experiment 4). Sprinkler irrigation until 140 DPS resulted in a bright shell tint; however continuing sprinkler irrigation from that date until harvest (at 165 DPS) resulted in a darker pod tint (Table 8). These data further supported the notion that the last 3 weeks of growth constitute the critical period for the effect of soil wetness on final shell tint.
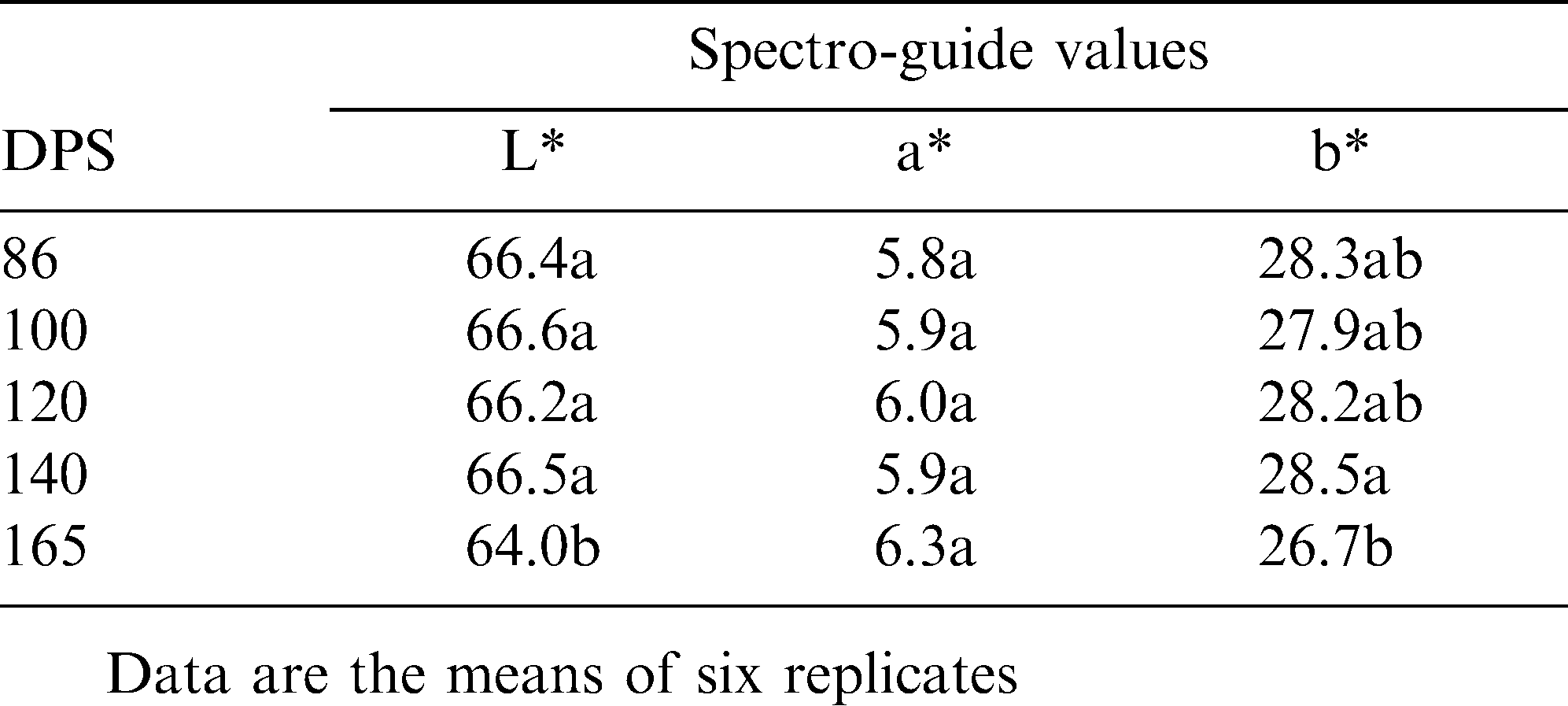
Determination of pod developmental stage most sensitive to soil wetness and loss of shell color by shifting from sprinkler irrigation to drip system (‘drip25’) at various time points after sowing — Field experiment 4.
Anatomy of Shell Development
During pod development, up until the pod maturation stage, the shell surface is smooth and white in color. Upon pod desiccation, the shell acquires its fibrous texture and yellowish color. It is suggested that at this latter stage (3–4 weeks before harvest), shell color may be influenced by soil wetness. Accordingly, pod wall development was histologically analyzed in an attempt to monitor morphological changes that may explain its texture and color during pod development (Figure 2). Although the dry tissue of the fully mature shell could not be viewed by the method used, the processes occurring at the pod wall prior to this stage could be seen. Starting from the seed-filling stage (Figure 2D), vascular bundles interlinked with the fibrous plate were seen to extend towards the surface of the pod wall, while the outer cell layers, which probably provide the pod's smooth texture and white color, appeared to be sloughing off (Figure 2F). Towards pod maturation, the fibrous material became part of the final pod wall/shell (Figure 2H) and it is suggested that this is the stage at which shell color is determined.
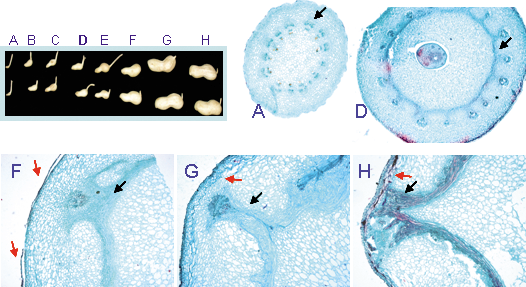
Anatomy of shell development. Cross sections were made of paraplast-embedded pods collected at various developmental stages (A–H, upper left panel). Sections were stained with Safranin-O/Fast green and viewed under a light microscope. The last stage of pod maturation — the dry shell — could not be processed by this technique, and section H therefore represents one stage prior to this. Sections show the vascular bundles (A, arrow) becoming interlinked by lateral connections (D, arrow). Cells elongate to form a fibrous plate (F, black arrow). From this plate, fibrous connections extend outward to the vascular bundle (G, black arrow). The mature pod wall consists mainly of the fibrous tissue and the vascular ramification (H, black arrow). Note the changes occurring in the outermost layers of the pod wall, implying sloughing of cells at the surface (red arrows).
Discussion
The data from this study show the involvement of soil type in determining final peanut shell color at maturity. Two main mechanisms were identified: absorption of soil material to, and loss of bright color from the shell surface. It is suggested that soil rich in organic matter will result in undesirable darker shells; however, this effect is dependent on the wetness of the soil around the pods and the developmental stage at which the pod is exposed to it.
In practice (in Israel), peanuts are harvested at around 165 DPS. The plant is pulled out of the soil and pods are allowed to air-dry under foliage cover for 10 to 14 days. The pod-drying conditions, e.g. air temperature and light exposure, may also affect shell tint; however, in the experiments described here, these conditions were kept the same between treatments.
High wetness resulted in loss of the desirable bright-yellow color of the shell. This is probably due to water-soluble soil components that adsorb to the shell surface, decreasing the shell's brightness. Likewise, high wetness in the pod-development zone may result in the dissolution of water-soluble shell-surface components, further reducing the yellowish brightness.
Accordingly, pods with an undesirable dark tint were obtained when the plant was irrigated by sprinklers or a drip-irrigation system placed above the pod-development zone (‘drip5’). Such conditions may result in increased soil wetness at the pod-development zone and ‘washing’ of the pods by the water flowing into the root zone. When water was applied directly to the root zone (‘drip25’), the pod-development zone was kept wet by capillary water, enough to enable normal development of the pods while preventing loss of shell color.
The difference in shell tint as a result of irrigation method was used to establish the developmental stage at which the final shell color is determined. A change from ‘drip5’ irrigation to ‘drip25’ at 100 DPS and the subsequent experiment in which sprinkler irrigation was changed to ‘drip25’ irrigation at various time points post-sowing established the last 3 weeks of growth as the critical period during which the final pod color is determined and the wetness of the soil near the pods needs to be controlled, respectively. It should be noted that although peanuts are indeterminate plants, the mature, marketable pods harvested at 165 DPS are at a similar developmental stage, and therefore similarly affected by soil wetness.
Field and histological observations (Figure 2) indicated that during pod growth/seed filling, the outer cell layers of its wall slough off and the immature shell remains white in color. Towards the end of growth, the layers that will make up the surface of the mature pod are exposed and the mature shell is formed. It is assumed that the effect of soil wetness on shell color occurs at this stage of pod-wall maturation.
Microscopic examination of the pod wall's surface at various stages of ‘White Spanish’ pod development detected root-hair-like epidermal outgrowths when the pod started to form (R3 stage, based on Boote, 1982), which disappeared from late R6 (full seed) to R8 (harvest maturity) (Webb and Hansen, 1989). Although the Hanoch cultivar used in the present study does not produce pod hairs, both cultivars demonstrate sloughing of the epidermal layers on the shell surface towards the stage of pod maturity (Webb and Hansen, 1989; Wissuwa and Ae, 1999, 2001). The subepidermal fibrous layers and the outer corky cells are exposed to the soil and probably interact with the pod's immediate microenvironment, affecting final shell tint.
The accumulated data suggest that soil wetness in the pod-development zone should be controlled during its final period growth to maintain a low level of wetness. This can be achieved by reducing the amount of water supplied by sprinklers or using a subsurface drip system placed between the pod zone and the root zone.
Acknowledgements
A contribution of ARO, the Volcani Center No. 127/2009, and Israel Peanuts Production and Marketing Board.
Literature Cited
Boote K. J. 1982 Growth stages of peanut (Arachis hypogaea L.). Peanut Sci 9 : 35 – 40 .
Chamarthy S. , Seo C. W. , and Marshall W. E. 2001 Adsorption of selected toxic metals by modified peanut shells. J. Chem. Technol. Biotechnol 76 : 593 – 597 .
Colwell W. E. , Brady N. C. , and Piland J. R. 1945 Composition of peanut shells of filled and unfilled fruits as affected by fertilizer treatments. J. Am. Soc. Agron 37 : 792 – 805 .
Henderson R. W. , Lightsey G. R. , and Poonawala N. A. 1977 Competitive adsorption of metal ions from solutions by low cost organic materials. Bull. Environ. Contam. Toxicol 18 : 340 – 344 .
Johansen D. A. 1940 Plant Microtechniques McGraw-Hill Book Company, Inc New York .
Johnson C. M. , Stout P. R. , Beyer J. C. , and Carlson A. B. 1957 Comparative chlorine requirements of different species. Plant and Soil 8 : 337 – 353 .
Periasamy K. and Sampoornam C. 1984 The morphology and anatomy of ovule and fruit development in Arachis hypogaea L. Ann. Bot 53 : 399 – 411 .
Pickett T. A. 1950 Composition of developing peanut seeds. Plant Physiol 25 : 210 – 224 .
Ruzin S. E. 1999 Plant Microtechnique and Microscopy Oxford University Press New York .
Wartelle L. H. and Marshall W. E. 2000 Citric acid modified agricultural by-products as copper ion adsorbents. Adv. Environ. Res 4 : 1 – 7 .
Webb A. J. and Hansen A. P. 1989 Histological changes of the peanut Arachis hypogaea gynophore and fruit surface during development, and their potential significance for nutrient uptake. Ann. Bot 64 : 351 – 357 .
Wilson K. , Yang H. , Seo C. W. , and Marshall W. E. 2006 Select metal adsorption by activated carbon made from peanut shells. Bioresource Technol 97 : 2266 – 2270 .
Wissuwa M. and Ae N. 1999 Genotypic variation for phosphorus uptake from hardly soluble iron- phosphate in groundnut (Arachis hypogaea L.). Plant and Soil 206 : 163 – 171 .
Wissuwa M. and Ae N. 2001 Genotypic differences in the presence of hairs on roots and gynophores of peanuts (Arachis hypogaea L.) and their significance for phosphorus uptake. J. Exp. Bot 52 : 1703 – 1710 .
Xu H. , Annis S. , Linz J. , and Trail F. 2000 Infection and colonization of peanut pods by Aspergillus parasiticus and the expression of the aflatoxin biosynthetic gene, nor-1, in infection hyphae. Physiol. Mol. Plant Pathol 56 : 185 – 196 .