

Introduction
Georgia continued to lead the US in peanut production with over 277,000 ha in 2008, and was second in cotton with over 384,000 ha (USDA-ARS, 2008). Since commercial introduction in 1997, glyphosate-tolerant cotton has rapidly been incorporated into most of the production across the southeast. In Georgia greater than 86% of the cotton ha was planted to one cultivar in 2008, Deltapine DP 555 BG/RR® (USDA-AMS, 2008). However, due to regulatory mandate, single gene Bollgard technology expired September 30, 2009 (Shurley et al., 2009). This regulation will end the use of Deltapine DP 555 BR/RR®. Glyphosate can only be postemergence directed applied to Roundup Ready® cultivars after reaching the four-leaf growth stage, thus limiting glyphosate use (May et al., 2004). Other cultivars, such as the second generation of glyphosate resistant Flex® (Murdock and Mullins, 2006) and Glytol® cotton (Trolinder et al., 2008), will replace DP 555 BR/RR®. One advantage of Flex® and Glytol® cotton cultivars is that producers may potentially apply glyphosate postemergence topical (POST) at any time during the growing season, up to just prior to harvest. This could lead to potential increase in glyphosate usage later during the growing season. As Georgia produces significant ha of cotton and peanut, the potential for glyphosate applied to peanut in error will likely increase.
Glyphosate drift and missapplication by tank contamination has been noted in many crops due to increased use in glyphosate resistant crops. Lassiter et al. (2007) reviewed the physiological effects of glyphosate for various non-glyphosate tolerant crops including rice (Oryza sativa L.), corn (Zea mays L.), cotton (Gossypium hirsutum L.), and soybean [Glycine max (L.) Merr.] noting that dependent on timing and rate of application, crops were either destroyed, severely injured reducing yield and quality, or exhibited little to no response. Glyphosate was marketed as a plant growth regulator in peanut (Colvin et al., 1990), but use was discontinued for unknown reasons. Lassiter et al. (2007) evaluated the Virginia peanut cultivar NC-12C (Isleib et al., 1997) to glyphosate at 9 to 1120 g ae/ha applied twice in the growing season to simulate drift. Glyphosate was applied at four weeks after planting and just prior to flowering. For this study, glyphosate at 280, 560, and 1,120 g/ha caused peanut injury and pod yield reduction. When peanut plant mass ranged from 10 to 15 cm in diameter and treated with glyphosate, a dose response ranging from no injury to severe stunting occurred for rates of glyphosate ranging from 9 to 1120 g/ha. When applied at doses up to 140 g/ha, peanut recovered and yield was not affected. Lassiter et al. (2007) evaluated peanut response to glyphosate in the vegetative stages of growth, making the second application prior to flowering, but they did not report the peanut growth stage (Boote, 1982) at time of application.
Glyphosate applied POST to Roundup Ready® cotton in reproductive stages of development (flowering) has reduced pollen production, caused formation of non-dehiscent anthers, and less viable pollen grains (Yasuor et al., 2007). Glyphosate applied to glyphosate resistant corn after the V6 stage of growth reduced pollen viability, although did not affect corn yield, which was attributed to the cross pollination ability of this crop (Thomas et al., 2004). Glyphosate applied during the reproductive stage of plant development is translocated and accumulated in metabolic sinks, such as reproductive tissues (Pline-Srnic, 2005). Wheat (Triticum aestivum L.) treated with glyphosate at various stages of reproduction development reduced seed kernel weight and lowered germination as compared to nontreated controls (Yenish and Young, 2000), with similar reports for Italian ryegrass (Lolium multiflorum Lam.) (Steadman et al., 2006). No information about glyphosate affects on peanut seed germination and vigor are reported. However, since peanut is primarily self-pollinated (Moss and Rao, 1995), there are potential negative affects to pollen, which could result in reduced seed viability, as observed in other species.
Previous research has noted Virginia type peanut tolerance to glyphosate will vary with the vegetative stage of growth and development (Lassiter et al., 2007). The effect of glyphosate applied to runner type peanut cultivars in reproductive stages of development is unknown. Thus, studies were conducted to evaluate glyphosate applied to runner type peanut at various reproductive stages of development to glyphosate for physiological effects on yield, seed development, and seed viability.
Materials and Methods
Experiments were conducted during 2006 and 2007 at the Southwest University of Georgia Research Station at Plains, and at the Ponder Research Farm located near Ty Ty Georgia in different areas of the same field. Soil for Plains was a Faceville sandy loam (clayey, kaolinitic, thermic, Typic Kandiudults) with 71% sand, 13% silt, and 16% clay. Soil for the Ty Ty site was a Tifton loamy sand (fine-loamy, kaolinitic, thermic Plinthic Kandiudults). Organic matter and pH were 1.0 to 1.5% and 6.0 to 6.5, respectively.
Peanut cultivars planted were GA-02C (Branch, 2003) at Ty Ty and AP-3 (Gorbet, 2007) at Plains. All seed were planted 4 cm deep at 20 seed row m−1 using a vacuum air planter. Planting date, herbicide application dates, harvest dates, and other parameters are presented in Table 1. Individual plots were two rows 91 cm wide by 7.6 m long at Ty Ty and two rows 91 cm wide by 9.1 m long at Plains. All experiments were three by six factorial treatment arrangements in a randomized complete block design (Gomez and Gomez, 1984) with four replications.
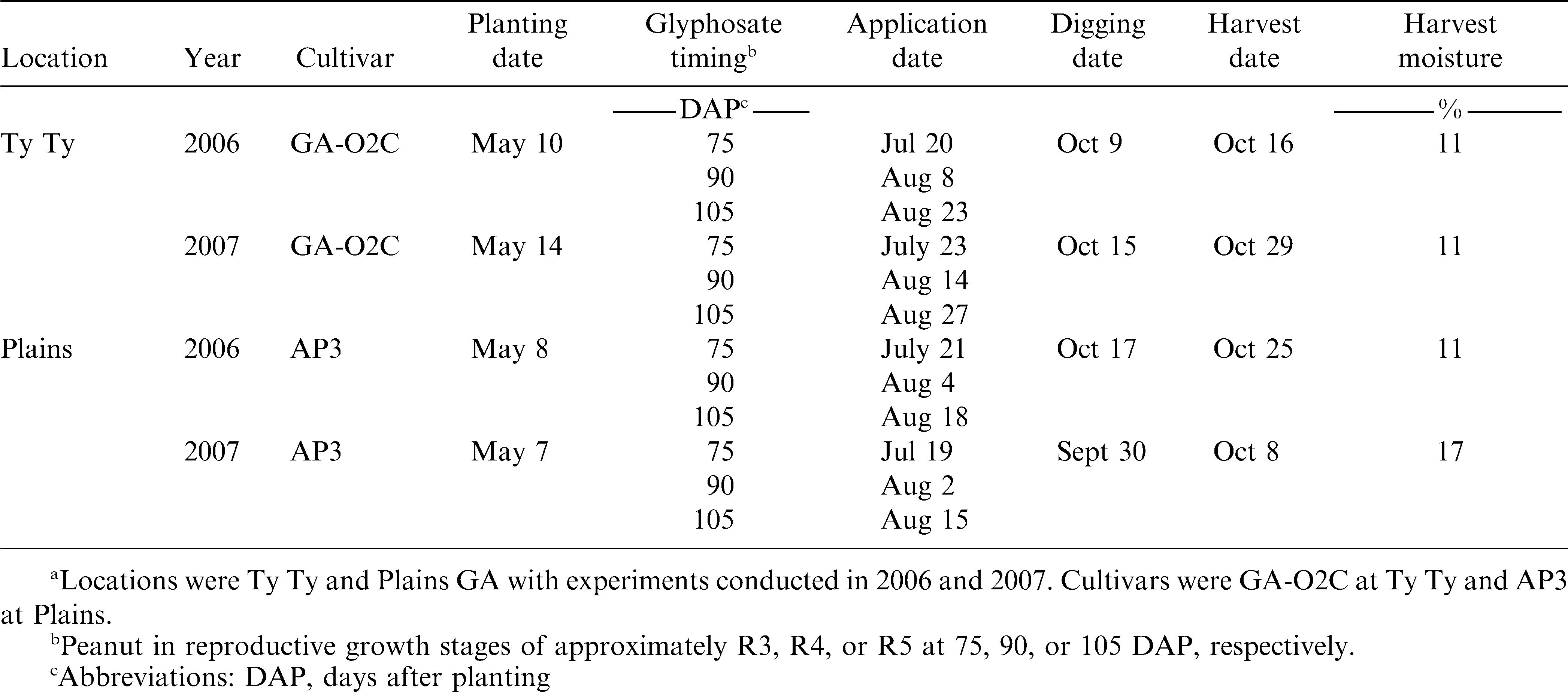
Test parameters for effect of glyphosate on runner type peanut in Georgia.a
Glyphostae rates evaluated were 80, 160, 240, 320, and 470 g ae/ha, and included a nontreated control for each application timing. These rates corresponded to 8, 17, 25, 33, and 50% of the recommend field application rates for glyphosate. Treatments were applied at 75, 90, or 105 days after planting (DAP) or when peanut was approximately in R3, R4, or R5 growth stages (Boote, 1982), respectively (Table 1). All experiments were maintained weed-free by using traditional weed control methods with the residual herbicides flumioxazin (105 g/ha), diclosulam (53 g/ha), and/or imazapic (71 g/ha) and hand-weeding. No other herbicides were applied. All glyphosate treatments were applied with a CO2-pressurized backpack sprayer calibrated to deliver 140 L/ha at 165 kPa in order to prevent mechanical damage.
Peanut was dug and inverted based on mesocarp pod color (Williams and Drexler, 1981). At digging, peanut seed samples were taken by randomly hand-harvesting 100 pods from each plot and stored for later evaluation. Field plots were harvested 10 to 20 d later with conventional harvesting equipment.
Harvested pods were hand shelled, allowed to air dry to 9% moisture at ambient temperature, and peanut seed mass determined. These same seed were used for germination testing. Seed were not treated with a fungicide. Peanut seed for each plot were randomly distributed on germination paper, which was placed in a 100 by 15 mm Petri dish. Ten seed were placed in each Petri dish followed by the addition of 10 ml of distilled water, and placed into a growth chamber set at a constant 25 C. Germination testing and counts were conducted according to official seed testing criteria for peanut (AOSA, 2000). Germination experiments were repeated in time for each field study.
All data were subjected to analysis of variance appropriate for the three (application timings) by six (herbicide) factorial treatment arrangement. Analysis of variance procedures were conducted with the MIXED procedure in SAS. Years and locations were regarded as random effects.
Results and Discussion
The two-way interactions between DAP and glyphosate rate was not significant for any variable (Table 2). Analysis of variance indicated DAP was significant for seed mass but no other variable. However, data for the main effects for glyphosate rate were significant for yield in kg/ha and kernel mass in mg/seed. Data for glyphosate rate was combined for analysis across DAP for all variables (Table 3). Data for DAP was also combined for analysis across glyphosate rate and presented or all variables (Table 4). Analysis of data indicated that rate of glyphosate was more important than the timing of application at 75, 90, or 105 DAP for peanut pod yield.
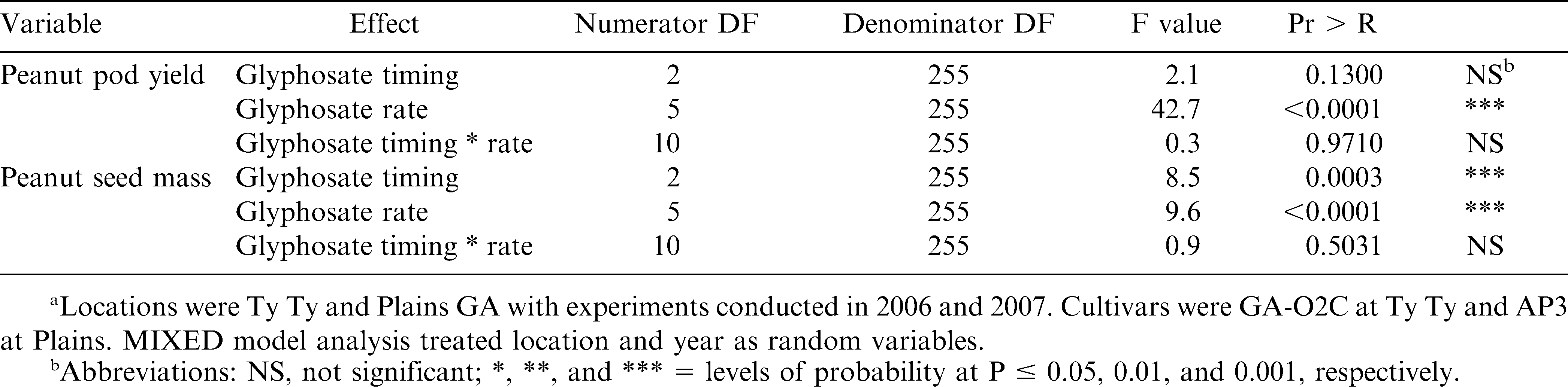
Analysis of variance for peanut pod yield and seed mass for glyphosate affects on runner type peanut in Georgia.a
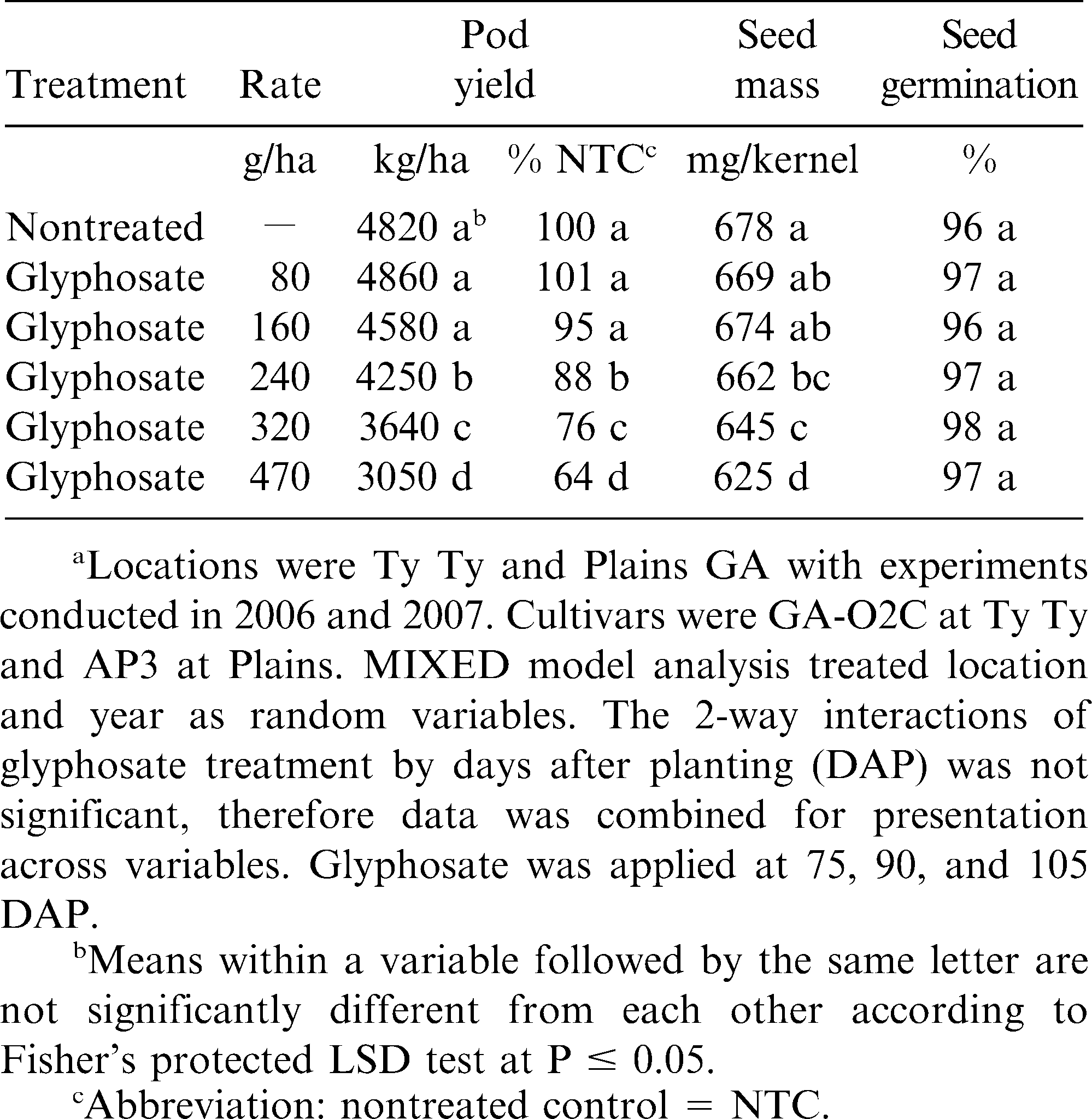
Glyphosate affects on runner type peanut pod yield, seed mass, and germination in Georgia.a
General observations indicated that peanut was tolerant to glyphosate at low doses, but did exhibit stunting and chlorosis (data not shown). Injury to peanut included leaf drop and some stand loss. Observations taken prior to peanut harvest indicated that peanut did not recover from injury for glyphosate doses of 240 g/ha and greater.
Peanut pod yield was reduced with increased glyphosate rate (Table 3). Glyphosate at 80 g/ha did not affect peanut, with yield equal to of the nontreated control. Glyphosate can have a plant growth regulative effect for peanut (Lassiter et al., 2007; Colvin et al., 1990). Although not different, glyphosate applied at 160 g/h reduced yield to 95% of the nontreated control. There were significant decreases to 88, 76, and 64% of peanut pod yield, compared to the nontreated control, for glyphosate at 240, 320, and 470 g/ha, respectively. Lassiter et al. (2007) evaluated shikimic acid accumulation in peanut and reported that low glyphosate rates (9 to 70 g/ha) did not increase shikimic acid levels as compared to nontreated controls. These data further dictate the need for producers to be diligent about misapplications of glyphosate on peanut. Although timing of glyphosate application was not significant, there was a trend for greater peanut pod yield reduction at 75 DAP (Table 4).
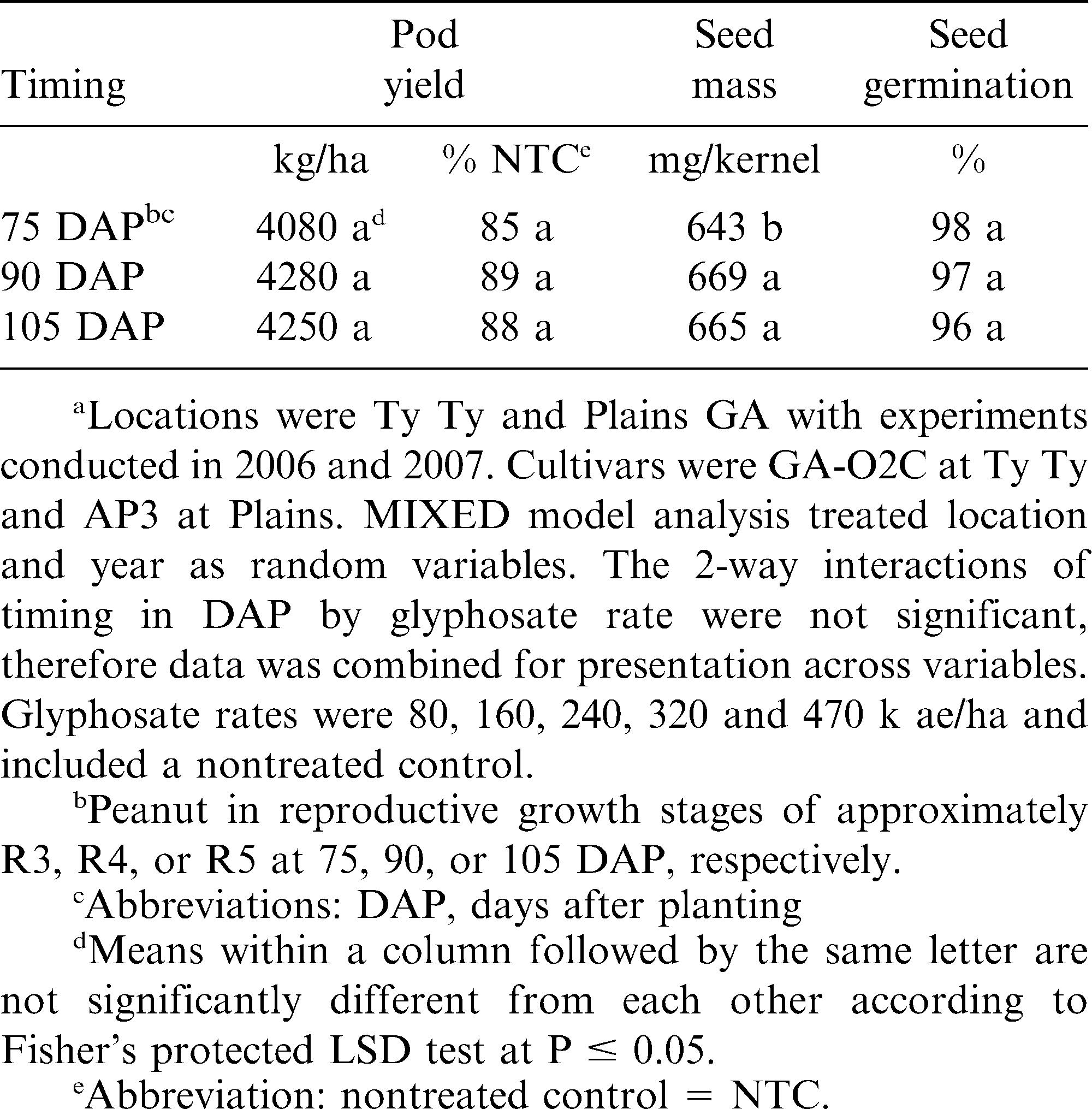
Timing affects of glyphosate on runner type peanut pod yield, seed mass, and germination in Georgia.a
Peanut seed mass was reflective of pod yield with significant mass reduction for glyphosate at 240 g/ha and greater (Table 3). In contrast, glyphosate at 80 or 160 g/ha did not reduce kernel mass as compared to the nontreated control. Winter wheat kernel mass reductions of 14% from glyphosate at 140 g ai/ha have been reported when applied at flowering (Roider et al., 2007). Peanut seed mass was reduced to 640 mg/kernel by glyphosate at 75 DAP as compared to the 90 and 105 DAP treatments (Table 4). Peanut was predominately in the R3 reproductive stage of growth for 75 DAP timing. As previously noted, glyphosate primarily translocates to plant meristimatic tissues. At the R3 stage of peanut, pod and seed are beginning to develop (Boote, 1982) and would thus be a major sink for glyphosate, although this has not been confirmed. While applications at the R4 and R5 stages of growth occurred when peanut was entering full pod to beginning seed, respectively. Previous research has indicated that glyphosate was translocated through chains of purple nutsedge tubers (Chase and Appleby, 1979; Elmasry and Rehm, 1977; Zandstra and Nishimoto, 1977) as well as other species (Duke et al., 2003). Thus, it is theorized that glyphosate would be translocated to peanut pods and seed as they develop.
Seed germination for all glyphosate rates did not reduce peanut germination as compared to the nontreated controls (Table 3). Although timing of glyphosate application did reduce peanut kernel mass for the 75 DAP treatment compared to other treatment timings, there was no adverse effect on peanut seed germination for any timing of application (Table 4). Previous research for glyphosate applied to purple nutsedge indicated reduced tuber viability and production, and that it does not metabolize in the plant (Doll and Piedrahita, 1982; Zandstra et al., 1974). It is theorized that as a high oil legume crop, peanut is similar to soybean with respect to glyphosate dissipation, but little is known about the degradation of glyphosate in plants (Duke et al., 2003).
Conclusions
Peanut yield was reduced by glyphosate rate, independent of timing of application at 75, 90, or 105 DAP. At rates of 240 g/ha and greater, peanut exhibited reduced peanut pod yield as well as reduced seed kernel mass. However, seed germination was not negatively effected by any glyphosate rate or timing of application. These data indicate that peanut does have some level of glyphosate tolerance, but the exact mechanism of tolerance and why seed germination were not negatively affected is not understood. Given that other species have exhibited reduced seed germination after glyphosate exposure (Roider et al., 2007; Steadman et al., 2006; Yasour et al., 2007; Yenish et al., 2000) but not others (Duke et al., 2003), it is theorized that peanut may have some method of glyphosate metabolism or conjugation that has yet to be discovered. This is an area of potential future research.
Acknowledgements
The authors wish to thank Charlie Hilton and Aaron Wise for technical assistance with this research. The authors thank the Georgia Peanut Commission for partial funding and assistance with this research.
Literature Cited
[AOSA] Association of Official Seed Analysts Rules for seed testing 2000 Association of Official Seed Analysts Las Cruces, NM .
Boote K. J. 1982 Growth stages of peanut (Arachis hypogaea L.). Peanut Sci 9 : 35 – 40 .
Branch W. D. 2003 Registration of ‘Georgia-02C’ peanut. Crop Sci 43 : 1883 – 1884 .
Chase R. L. and Appleby A. P. 1979 Effects of humidity and moisture stress on glyphosate control of Cyperus rotundus L. Weed Res 19 : 241 – 246 .
Colvin D. L. , MacDonald G. E. , Shilling D. G. , Mossler M. A. , Kvien C. , Swan C. W. , and Wehtje G. R. 1990 Physiological and yield effects on peanut (Arachis hypogaea L.) from foliar applied yield enhancers. Proc. South. Weed Sci. Soc 43 : 109 .
Doll J. D. and Piedrahita W. 1982 Effect of glyphosate on the sprouting of Cyperus rotundus L. tubers. Weed Res 22 : 123 – 128 .
Duke S. O. , Rimando A. M. , Pace P. F. , Reddy K. N. , and Smedaj R. J. 2003 Isoflavone, glyphosate, and aminomethylphosphonic acid levels in seeds of glyphosate-treated, glyphosate-resistant soybean. Agric. Food Chem 51 : 340 – 344 .
Elmasry R. and Rehm S. 1977 Effects of growth-regulators and herbicides on purple nutsedge (Cyperus rotundus L).4. Effects of glyphosate and morphactin on viability of tubers. Zeitschrift Fur Acker Und Pflanzenbau-J. Agron. Crop Sci 144 : 259 – 267 .
Gorbet D. W. 2007 Registration of ‘AP-3’ Peanut. J. Plant Registrations 2 : 126 – 127 .
Gomez K. A. and Gomez A. A. 1984 Factorial experiments. pp. 89 – 91 In Statistical procedures for agricultural research. 2nd ed John Wiley & Sons New York .
Isleib T. G. , Rice P. W. , Bailey J. E. , Mozingo R. W. , and Pattee H. E. 1997 Registration of ‘NC 12C’ peanut. Crop Sci 37 : 1976 .
Lassiter B. L. , Burke I. C. , Thomas W. E. , Pline-Srnic W. A. , Jordan D. L. , Wilcut J. W. , and Wilkerson G. G. 2007 Yield and physiological response of peanut to glyphosate drift. Weed Technol 21 : 954 – 960 .
May O. L. , Culpepper A. S. , Cerny R. E. , Coots C. B. , Corkern C. B. , Cothren J. T. , Croon K. A. , Ferreira K. L. , Hart J. L. , Hayes R. M. , Huber S. A. , Martens A. B. , McCloskey W. B. , Oppenhuizen M. E. , Patterson M. G. , Reynolds D. B. , Shappley Z. W. , Subramani J. , Witten T. K. , York A. C. , and Mullinix B. G. 2004 Transgenic cotton with improved resistance to glyphosate herbicide. Crop Sci 44 : 234 – 240 .
Moss J. P. and Rao V. R. 2005 The peanut – reproductive development to plant maturity. In Pattee H. E. and Stalker H. T. (eds.) Advances in Peanut Science Amer. Peanut Res. Educ. Soc., Inc., Stillwater, OK .
Murdock S. W. and Mullins W. 2006 Roundup Ready® Flex cotton - 2006 launch. Proc. Beltwide Cotton Conf, National Cotton Council, Memphis, TN. Online at http://www.cotton.org/beltwide/proceedings/2005–2009/data/conferences/2006/pdfs/2300–2301.pdf. (Verified June 6, 2009).
Pline-Srnic W. 2005 Technical performance of some commercial glyphosate-resistant crops. Pest Manag. Sci 61 : 225 – 234 .
Roider C. A. , Jones C. A. , Harrison S. A. , and Griffin J. L. 2007 Wheat response to simulated glyphosate drift. Weed Technol 21 : 1010 – 1015 .
Shurley W. D. , Day J. L. , Roberts P. R. , and Culpepper A. S. 2009 Expiration of single-gene Bollgard® technology: analysis of alternatives available to Georgia cotton producers. Proc. Beltwide Cotton Conf, National Cotton Council, Memphis, TN. Online at http://www.cotton.org/beltwide/proceedings/2005–2009/data/conferences/2009/papers/9507.pdf (verified June 6, 2009).
Steadman K. J. , Eaton D. M. , Plummer J. A. , Ferris D. G. , and Powles S. B. 2006 Late-season non-selective herbicide application reduces Lolium rigidum seed numbers, seed viability, and seedling fitness. Australian J. Agri. Res 57 : 133 – 141 .
Thomas W. E. , Pline-Srnic W. A. , Thomas J. F. , Edmisten K. L. , Wells R. , and Wilcut J. W. 2004 Glyphosate negatively affects pollen viability but not pollination and seed set in glyphosate-resistant corn. Weed Sci 52 : 725 – 734 .
Trolinder L. , Ellis J. M. , Holloway J. , and Baker S. 2008 GLYTOL cotton – new herbicide tolerant cotton from Bayer CropScience. Proc. Beltwide Cotton Conf, National Cotton Council, Memphis, TN. Online at http://www.cotton.org/beltwide/proceedings/2005–2009/data/conferences/2008/papers/8013.pdf (verified June 6, 2009).
United States Department of Agriculture - Agricultural Marketing Service - Cotton Program (USDA-AMS) 2009 Cotton Varieties Planted 2009 Crop. Memphis, TN. Online at http://www.cottoninc.com/CropQualitySummary/CropQualitySummaryWeekly/MOST-POPULAR-VARIETIES-FOR-2009.PDF (verified April 7, 2010).
United States Department of Agriculture-National Agricultural Statistics Service (USDA-NASS) 2008 Agricultural Chemical Usage Estimates for Field Crops. http://www.nass.usda.gov. (verified June 6, 2009).
United States Department of Agriculture-National Agricultural Statistics Service (USDA-NASS) 2008 Crop Production Annual Summary. http://www.nass.usda.gov. (verified June 29, 2009).
Williams E. J. and Drexler J. S. 1981 A non-destructive method for determining peanut pod maturity. Peanut Sci 8 : 134 – 141 .
Yasuor H. , Riov J. , and Rubin B. 2007 Glyphosate-induced male sterility in glyphosate-resistant cotton (Gossypium hirsutum L.) is associated with inhibition of anther dehiscence and reduced pollen viability. Crop Prot 26 : 363 – 369 .
Yenish J. P. and Young F. L. 2000 Effect of preharvest glyphosate application on seed and seedling quality of spring wheat (Triticum aestivum). Weed Technol 14 : 212 – 217 .
Zandstra B. H. and Nishimoto R. K. 1977 Movement and activity of glyphosate in purple nutsedge. Weed Sci 25 : 268 – 274 .
Zandstra B. H. , Teo C. K. H. , and Nishimoto R. K. 1974 Response of purple nutsedge to repeated applications of glyphosphate. Weed Sci 22 : 230 – 232 .
Notes
- 1Assoc. Prof. and 2Prof., Dept. of Crop and Soil Sci., respectively, The University of Georgia, Tifton Campus, P.O. Box 748, 115 Coastal Way, Tifton, GA 31793. [^] *Corresponding author (email: tgrey@uga.edu).
Author Affiliations