

Introduction
Sclerotinia blight, caused by the fungus Sclerotinia minor Jagger, was first reported on peanut (Arachis hypogaea L.) in the United States in Virginia during 1971 (23) and was identified in Oklahoma in 1972 (36). The disease now is endemic to peanut production areas of North Carolina, Virginia, Oklahoma, and Texas where the pathogen persists for extended periods in soil as sclerotia. Once the plant canopy develops and soil moisture is high, sclerotia near the soil surface germinate to produce mycelia that infects nearby plant parts (26, 35). Stems, pegs, leaves, and pods are susceptible to infection (35). Stem lesions kill individual branches and eventually entire plants as the mycelium spreads within the plant canopy during periods of cool and wet conditions that favor disease development. Yield loss is generally proportional to the percentage of plants affected and results primarily from stem and peg decay that causes pods to become detached from plants during digging.
Management of Sclerotinia blight has relied on an integrated program of cultural practices, partially resistant cultivars, and fungicide programs (25). Crop rotation is effective for reducing sclerotial build-up, but has little practical value for problem fields with a high sclerotial density because of sclerotial persistence. Sanitation practices are directed at limiting the spread of the fungus by preventing its introduction into new fields. The fungicides fluazinam and boscalid provide good disease control when applied preventively or at the first sign of disease (9, 10, 34). However, the cost per application of these fungicides ranges from $100 to $150/ha depending on the rate used. The use of cultivars with genetic resistance to Sclerotinia blight is a more economical approach to disease management.
In the evaluation of peanut cultivars and breeding lines for resistance to Sclerotinia blight, various levels of partial resistance have been identified in spanish, runner, and virginia market types (1, 4, 6, 20, 24). Partial resistance in these market types has two forms. Physiologic resistance is expressed as reduced lesion expansion over time as measured on inoculated seedlings or detached plant parts following incubation under artificial conditions favorable for disease development (1, 4, 15, 20, 21). Architectural or morphological resistance is expressed in some genotypes with an erect growth habit and sparse plant canopy that results in reduced contact of main and lateral stems with mycelium from the germinating sclerotia or adjacent infections; and/or reduced periods of microclimate favorable for disease development compared to spreading genotypes with a dense canopy (1, 4, 6). Architectural resistance is apparent when genotypes are grown without artificial inoculation in an infested field. Based on local experiences in peanut production areas in the southwestern U.S. where runner and spanish market types have predominated, both forms of partial resistance are effective. Tamspan 90 (32) is a spanish cultivar with both architectural and physiologic partial resistance (1). Southwest Runner (22) is a runner-type cultivar with sparsely arranged large leaves and a prominently erect main stem that is susceptible in detached stem and seedling inoculations in the greenhouse. Both Tamspan 90 and Southwest Runner are consistently among the most resistant cultivars in field evaluations. Ratings from assays designed to identify physiologic resistance on whole or detached plant parts are generally not well correlated with field screenings (1, 15, 20, 21)
Tamspan 90 was grown on nearly all of the problem fields in Oklahoma during the 1990's, but susceptibility to web blotch (caused by Phoma arachidicola Marassas, G.D. Pauer & Boerema) and reduced demand for spanish peanut have contributed to the increase in acreage planted to runner cultivars. While Southwest Runner is partially resistant to Sclerotinia blight, it was never accepted by the shelling industry because of its small seed size. Alternatively, runner cultivars from the Texas A&M breeding program such as Tamrun 96 (33), Tamrun 98 (30), Tamrun OL01 (28), and Tamrun OL02 (29) have been released and grown in problem fields. The level of partial resistance in these cultivars is generally intermediate between the susceptible cultivars Florunner and Okrun (3) and the resistant cultivars Tamspan 90 and Southwest Runner, and is not sufficient to preclude the need for fungicide applications (8, 10). Except for some trials with Tamspan 90 and Southwest Runner, the effects of partial resistance and fluazinam application on yield have been additive, indicating a need for improving the level of resistance to Sclerotinia blight.
Germplasm collections represent additional potential sources of genetic resistance to Sclerotinia blight and other diseases. The USDA collection of peanut (Arachis hypogea) germplasm is comprised of over 9,000 accessions (16). Because of the large number of accessions in this and other germplasm collections, a core collection approach to increasing the efficiency of collection evaluation and trait utilization was proposed (12). The core collection approach to germplasm evaluation is a two-stage process that first involves screening a reduced number of accessions and using this information to identify clusters of accessions in the entire collection with increased probability of identifying the desired characteristic (16). The USDA collection of A. hypogea germplasm was the first to have a working core collection. The core collection is comprised of 831 entries selected at random (1%), at random by country of origin (26%), and by multivariate clustering of phenotypic descriptor information (73%) to minimize repetitiveness in the collection while representing the collection's genetic diversity (16,18). The peanut core collection has been previously evaluated for resistance to tomato spotted wilt virus (TSWV), Rhizoctonia limb rot, late leaf spot, peanut root knot nematode, preharvest aflatoxin contamination, early leaf spot, and Cylindrocladium black rot (2, 13, 16, 17, 19). To our knowledge, the core collection has not been previously evaluated for reaction to Sclerotinia blight. Therefore, the objective of this research was to evaluate the core collection in the field for reaction to Sclerotinia blight in hopes of identifying new sources of resistance to the disease. In the course of the evaluations, information also was obtained on entry reactions to pepper spot (caused by Leptosphaerulina crassiasca (Sechet) C.R. Jackson & D.K. Bell) and web blotch which became apparent during screenings. Brief excerpts of this research have been previously published (5, 11).
Materials and Methods
Field trials for evaluating core collection entries for reaction to Sclerotinia blight were conducted at the Caddo Research Station near Ft. Cobb, OK from 2001 to 2003 in fields containing a Meno fine sandy loam previously cropped to peanut and having a history of Sclerotinia blight. Soil samples from the fields were assayed by wet sieving and contained up to 3 sclerotia per 100 cc soil. The fields received 112 kg/ha of 18N-46P-0K granular fertilizer and were ripped, plowed, disked, and bedded prior to planting each year.
In 2001, 751 of the 831 accessions in the core collection were available. Because only 100 seeds of each entry were received, entries were planted in non-replicated plots. The herbicide pendimethalin was applied pre-plant incorporated at 0.55 kg/ha prior to planting on 23 May. Plots consisted of two, 3-m-long rows spaced 0.91 m apart. Metolaclor was applied pre-emergence at 0.71 kg/ha on 25 May for additional weed control. A tank mixture of propiconazole at 0.063 kg/ha and chlorothalonil at 0.84 kg/ha was applied on 2 Jul, 1 Aug, and 4 Sep for control of early leaf spot and other foliar diseases. Other cultural and pest management practices were applied according to extension service recommendations (14). Rainfall during the cropping period totaled 1.7 cm for June and July, 8.5 cm for August, 1.7 cm for September, and 0.5 cm for October. The field received 9 applications of water by sprinkler irrigation at 3.8 to 5.1 cm/application that totaled 43.2 cm from 27 Jun to 8 Oct. Incidence of Sclerotinia blight was assessed on 12 Sep and 11 Oct by counting the number of 15-cm row sections with symptoms and/or signs of disease. The counts were adjusted to the percentage of row length affected. Symptoms of foliar disease consistent with pepper spot became apparent in the trial and were evaluated on the same assessment dates by visually estimating the percentage of leaflets with symptoms which included defoliation. Entries with a nil to low (<10%) incidence of Sclerotinia blight and plant characteristics warranting further evaluation were marked, dug on 19 Oct, and dried in windrows for three days. Pods were harvested with a stationary plot thresher and dried to ca. 10% moisture prior to shelling and storage. Disease incidence data was obtained for 744 entries that had sufficiently viable seed to produce an adequate plant stand. Pearson correlation analysis was used to explore associations between incidences of Sclerotinia blight and pepper spot, and among disease incidence and the descriptors of maturity (1 = earliest, 6 = latest), growth habit (2 = most prostrate, to 6 = most erect), and plant size (1 = smallest, 6 = largest) using the CORR procedure of SAS (Statistical Analysis Systems, version 9.1, SAS Institute, Cary, NC). Frequency distributions using categories of Sclerotinia blight incidence (0, 1 to <10%, 10% to <25%, 25% to <50%, 50 to <75%, and 75% to 100%) were constructed using FREQ procedure of SAS. Chi square (X 2) estimates were used to test for deviation of observed frequencies from equal proportions among the five categories of disease incidence, and to compare frequency distributions among two-way comparisons of the maturity group and growth habit descriptors.
In 2002 and 2003, entries selected for further evaluation were planted in replicated trials at the Caddo Research Station. In 2002, 77 core collection entries were planted in a randomized incomplete block design along with reference cultivars that have variable reactions to Sclerotinia blight. The reference cultivars were Tamspan 90 (resistant), Tamrun 96 (moderately resistant), and Okrun (susceptible; 8,10). The number of replications for the core entries was two to four because sufficient quantities of seed were not produced in 2001 to plant four replications of all entries. The trial was planted on 8 May using the same cultural practices and plot dimensions as in 2001. Ethafluralin was preplant incorporated at 0.52 kg/ha for weed control. The herbicides 2,4DB at 0.22 kg/ha + bentazon at 0.56 kg/ha were applied post-emergence on 21 Jun, and 2,4DB at 0.22 kg/ha + crop oil at 1.17 l/ha were applied post-emergence on 8 Jul for additional weed control. Foliar diseases were controlled with the tank mixture of propiconazole + chlorothalonil described above and applied on 2 Jul and 22 Jul, and with tebuconazole at 0.23 kg/ha applied on 5 Aug and 19 Aug. Tebuconazole also has activity on the soilborne diseases stem rot (caused by Sclerotium rolfsii) and limb rot (caused by Rhizoctonia solani), but not Sclerotinia blight. Rainfall during the cropping period totaled 11.1 cm in June, 4.6 cm in July, 3.5 cm in August, 6.5 cm in September, and 15.1 cm in October. Sprinkler irrigation was applied 8 times from 15 Jul to 30 Sep at 5 cm water per application. Incidences of Sclerotinia blight and pepper spot were evaluated as described above on 20 Sep and 30 Oct. Plots were dug on 13 Nov and harvested as previously described prior to weighing.
In 2003, 62 core collection entries were planted in a randomized complete block design along with the same reference cultivars described above. The 15 entries that were screened in the replicated trial in 2002 but not included in 2003 were dropped because they had a high incidence (>25%) of Sclerotinia blight. The herbicide ethafluralin at 0.63 kg/ha was incorporated into the soil prior to planting on 20 May. Plots consisted of two 4.6 m-long rows spaced 0.91 m apart. Diclosulam was applied pre-emergence at 0.18 kg/ha for additional weed control on 22 May. Foliar diseases were controlled with the tank mixture of propiconazole and chlorothalonil on 30 Jun, 14 Jul, and 8 Sep; and foliar disease and stem rot were controlled with tebuconazole on 28 July, 12 Aug, and 21 Aug. Rainfall during the cropping period totaled 16.8 cm for June, 2.7 cm for July, 8.4 cm for August, 2.03 cm for September, and 2.3 cm for October. Plots received 12 applications of sprinkler irrigation that totaled 26.0 cm of water at 1.3 to 2.5 cm per application from 30 May to 26 Sep. Incidences of Sclerotinia blight and pepper spot were assessed as described above on 19 Sep and 10 Oct. Web blotch also developed in the plots and disease incidence was assessed in the same manner as for pepper spot. Plots were dug on 24 Oct and harvested 29 Oct as described above.
Data on final disease incidence and yield from the replicated trials in 2002 and 2003 were subjected to analysis of variance using the GLM procedure of SAS. The random effects of year and block and the fixed effects of entry were tested in a mixed model that evaluated the year, block (year), entry, and year*entry. Least square means were compared using t-tests produced from the PDIFF option of the LSMEANS statement in Proc GLM. Letters were assigned to the mean separation groupings using a freely available macro (27). Only significant differences (P ≤ 0.05) will be described below unless otherwise noted.
Results
In 2001 when entries of the core collection were planted in non-replicated plots, conditions did not favor early development of Sclerotinia blight. Rainfall was over 60% below normal (30-yr mean) from June through October. However, rainfall was 25% above normal for August and monthly average temperature was below normal for August through October which, along with irrigation, favored disease development. Sclerotinia blight was identified in the trial in September and reached severe levels by harvest. Susceptible check cultivars were not included in this trial, but disease incidence in some entries exceeded 90%. Pepper spot also appeared in the trial and exceeded an incidence of 70% for some entries.
There was considerable variation in the incidence of Sclerotinia blight just prior to harvest among the 744 entries evaluated (Figure 1) as the distribution of entries in the six disease incidence categories deviated from equal proportions (X 2 = 116.4, P < 0.01). Nearly 30% of the entries had <10% disease incidence while only 21% had 50% disease incidence or more. Incidence of Sclerotinia blight was positively correlated with maturity (r = 0.50, P < 0.01), negatively correlated with growth habit (r = −0.62, P < 0.01), but was not correlated with plant size (r = 0.03, P = 0.45). In comparing the distributions of incidence of Sclerotinia blight by maturity group, entries in maturity groups 1, 5 and 6 were not included because there were only 22 entries in group 1, only 5 entries in group 6, and no entries in group 5. The distributions of disease incidence categories differed between maturity groups 2 and 3 (X 2 = 66.6, P < 0.01), between maturity groups 2 and 4 (X 2 = 89.7, P < 0.01), and between maturity groups 3 and 4 (X 2 = 11.5, P = 0.04). Distributions of disease incidence categories were skewed towards low disease incidence for maturity group 2 and for high disease incidence for maturity group 4 (Figure 2). In maturity group 2, most (68%) of the entries had <25% disease incidence, while most (71%) of the entries in maturity group 4 had 25% or more disease incidence. In maturity group 3, about half of entries had less than 25% and the other half more than 25% disease incidence.
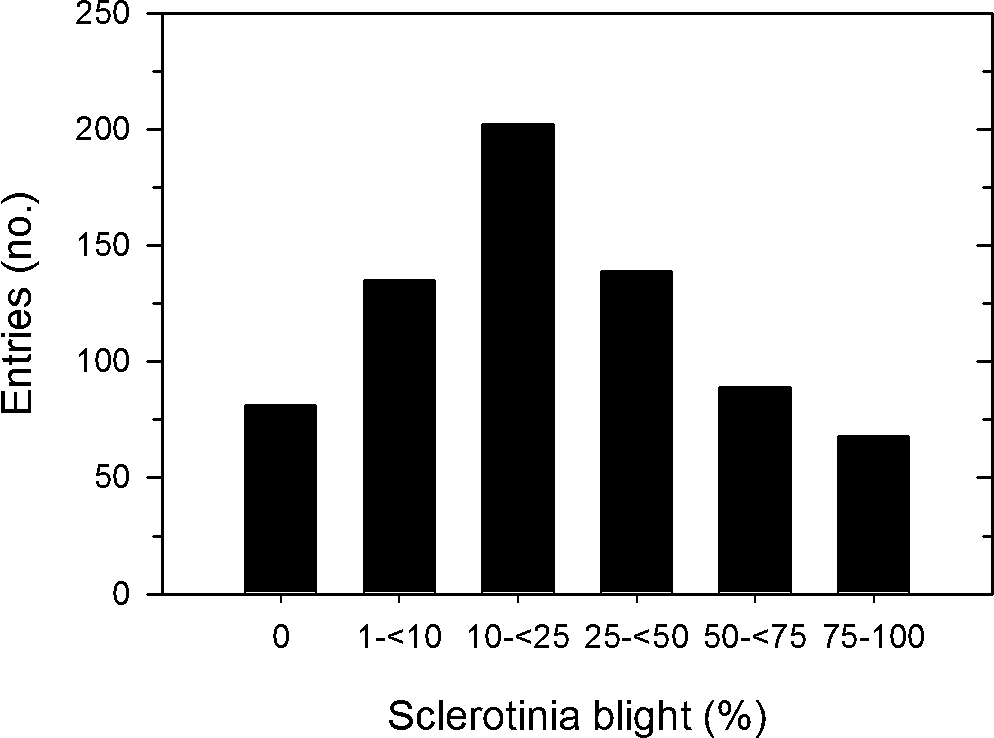
Frequency distribution of the field reaction to Sclerotinia blight of 744 entries from the peanut core collection at the Caddo Research Station in 2001.
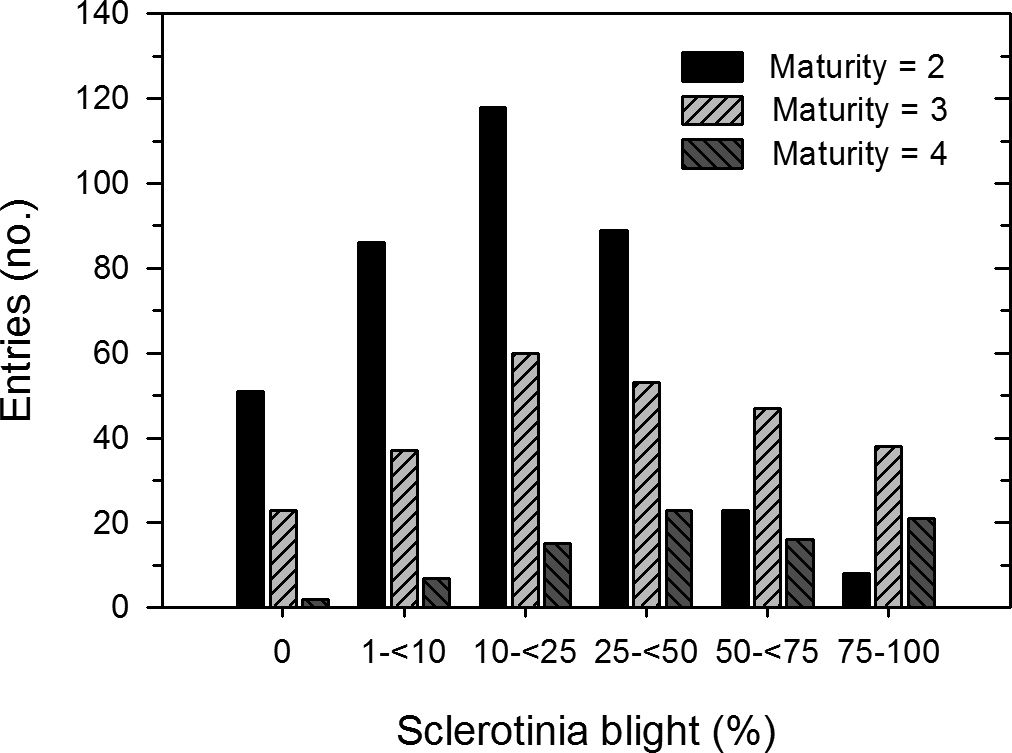
Frequency distribution of the field reaction to Sclerotinia blight of 744 entries from the peanut core collection at the Caddo Research Station in 2001 by maturity group (2 = early maturity, 4 = late maturity).
Frequency distributions of Sclerotinia blight incidence varied by plant growth habit (Figure 3). Growth habit 1 (most prostrate) and 6 (most erect) were omitted from the analysis because there were only 1 and 27 entries in these descriptor categories, respectively. The distributions of Sclerotinia blight incidence among all pairwise comparisons of the growth habit descriptors of 2 to 5 differed (P > X 2 < 0.001) except for growth habits 3 and 4 (X 2 = 6.6, P = 0.25). For growth habits 2 to 4, most (88%, 72% and 58%, respectively) of the entries were in the susceptible categories of 25% or more disease incidence. In growth habit 5, most (72%) of the entries were in the more resistant categories of <25% disease incidence.
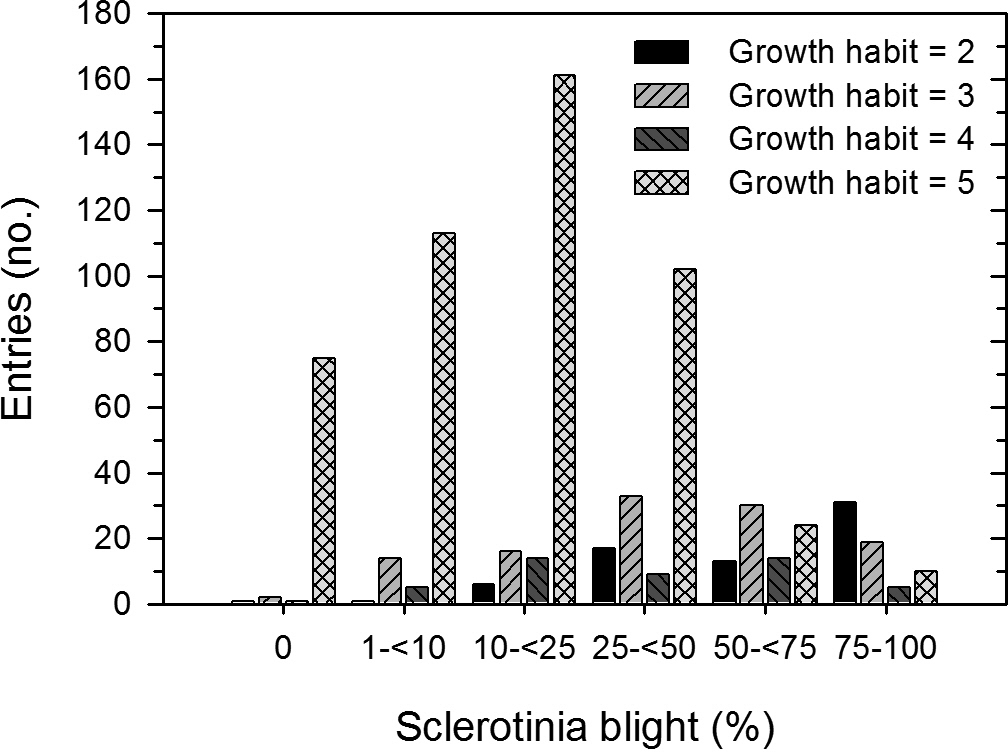
Frequency distribution of the field reaction to Sclerotinia blight of 744 entries of peanut core collection to Sclerotinia blight at the Caddo Research Station in 2001 by plant habit (2 = prostrate, 5 = upright).
The fungicide spray program applied for foliar diseases was highly effective against early leaf spot, but not against pepper spot which reached an incidence of up to 80% for the most susceptible entries. Although defoliation caused by pepper spot reached 40% for a few entries, most of the entries with pepper spot had no defoliation. There was considerable variation in the response of the 744 entries to pepper spot. There were 32% of the entries (n = 235) that had no disease and 18% (n = 131) that had a disease incidence of over 50% symptomatic leaves. The remaining 50% of the entries had from 5 to 50% disease incidence. Incidence of pepper spot was negatively correlated with maturity group (r = −0.55, P < 0.01), positively correlated with growth habit (r = 0.52, P < 0.01), and was not associated with plant size (r = 0.01, P = 0.70). Incidence of pepper spot was negatively correlated with incidence of Sclerotinia blight (r = −0.43, P < 0.01).
In 2002, weather conditions generally favored disease development as rainfall from June through October was 7% above normal and average daily temperature was below normal each month. However vine growth was not vigorous in the trial and the vines of most entries did not overlap between adjacent rows. Pepper spot was also severe in the trial. As a result, Sclerotinia blight did not appear until mid-September, about a month later than in adjacent trials on the research station. In 2003, rainfall was 15% below normal from June through October. However, rainfall in August was above normal and average daily temperature during September was 2.7 C below normal which favored disease development. Sclerotinia blight appeared in late August and reached severe levels by harvest. Pepper spot also developed in 2003, but the level of disease incidence was 50% less than in 2002. The cool temperatures in September also favored web blotch which reached moderate levels for some of the entries. In the analysis of variance, the effects of year, entry, and the year x entry were significant for incidence of Sclerotinia blight, pepper spot, and yield. Therefore, disease and yield data were presented by year.
In the replicated trials conducted in 2002 and 2003, Sclerotinia blight developed each year, but disease incidence was greater in 2003 compared to 2002 (Table 1). Of the reference cultivars, the susceptible cultivar Okrun had the highest disease incidence each year. The moderately resistant cultivar Tamrun 96 had about 50% less disease than Okrun in 2002, but the cultivars did not differ in 2003. The resistant spanish cultivar Tamspan 90 had less than 10% disease each year and was the most resistant cultivar. The core entries 329, 238, 466, 763, 599, 463, 227, 345, 249, 679, 326, and 457 were selected for further evaluation because of their low incidence of Sclerotinia blight in 2001. However, these entries had a moderate (26%) to high (46%) incidence of Sclerotinia blight in 2002 and were not included in the 2003 trial. These were all mid-maturing entries in maturity groups 3 and 4, that mostly had prostrate (growth habit = 2) to intermediate (growth habit = 3 to 4) growth habits. However, entries 466, 763, 249, and 599 had erect growth habits (growth habit = 5).
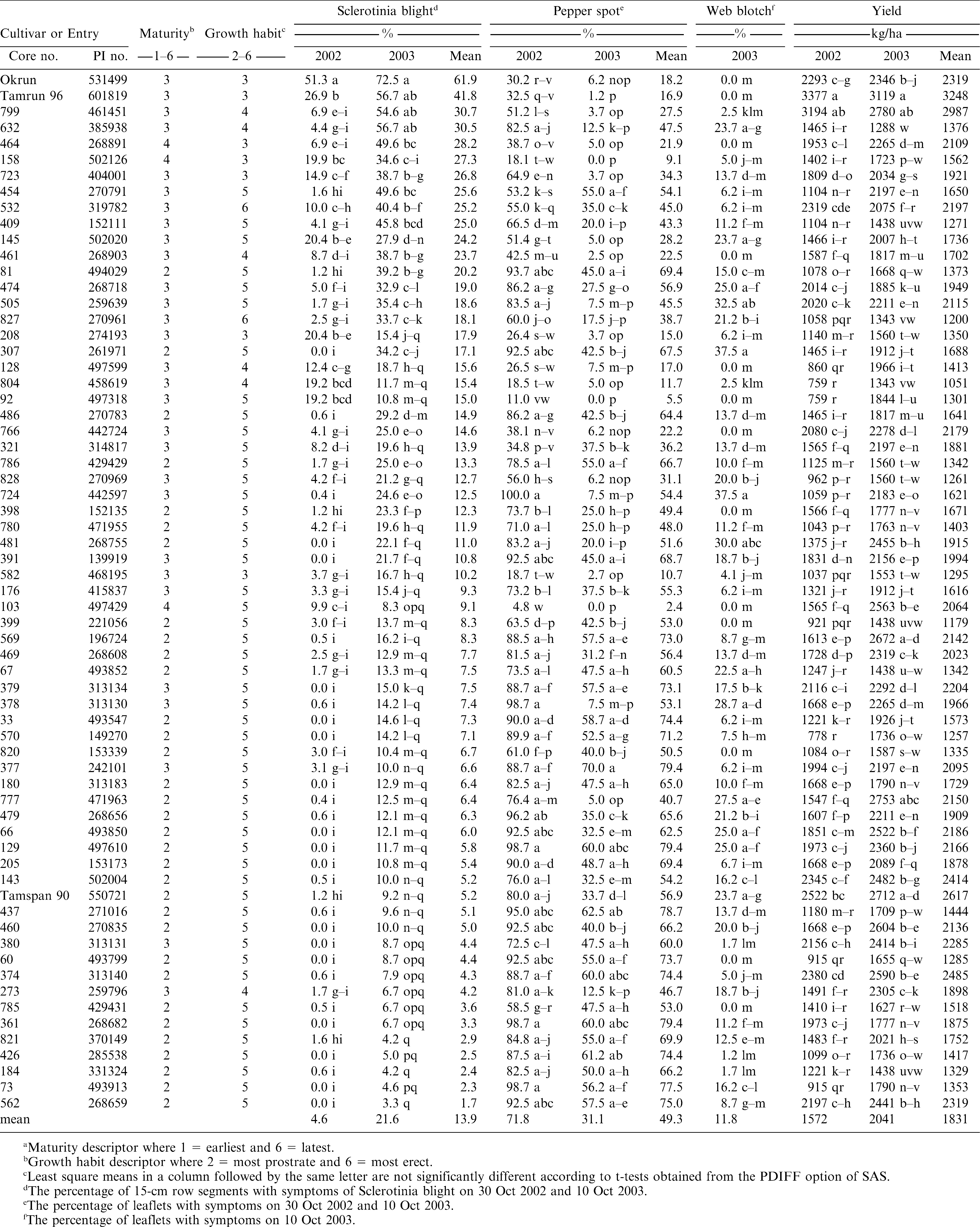
Least squares estimates of disease incidence and yield for entries of the peanut core collection and reference cultivars selected for response to Sclerotinia blight at the Caddo Research Station in 2002 and 2003.a
Among the entries grown both in 2002 and 2003 (Table 1), most of the entries were in maturity group 2 (n = 32), similar to Tamspan 90; or in group 3 (n = 27) similar to Okrun, and Tamrun 96. Only entries 464, 158, 103 were in the late maturing group 4. Most of the entries (N = 50) had an erect growth habit similar to Tamspan 90 (growth habit = 5). Only entries 532 and 827 had very erect growth habits (growth habit = 6). Entries 464, 158, 723, 208, and 582 had prostrate plant types (growth habit = 3) similar to Okrun and Tamrun 96. Entries 799, 632, 461, 128, 804, and 273 had intermediate growth habits (growth habit = 4). There were no entries with a very prostrate growth habit (growth habit = 2) similar to Georgia Green. All entries except 799 and 632 had disease incidence less than Okrun both years. Most entries (55 in 2002 and 46 in 2003) had disease incidence less than the moderately resistant Tamrun 96 and similar to the resistant cultivar Tamspan 90. In 2003 when disease incidence was highest, all of the 46 entries with resistant reactions comparable to Tamspan 90 had erect growth habits (growth habit = 5) except for entries 208 and 582 which had a prostrate growth habit (growth habit = 3); and entries 273, 128, and 804 which had an intermediate growth habit (growth habit = 4). Entries 454, 409, 145, 474, 81, 505, 307, and 486 had erect plant habits, but intermediate levels of Sclerotinia blight greater than Tamspan 90. Entries 827 and 532 were the only two entries with very erect growth habits (growth habit = 6), but had intermediate levels of Sclerotinia blight that were greater than Tamspan 90.
Fungicide programs used in 2002 and 2003 were highly effective against early leaf spot, but not pepper spot. Pepper spot reached an incidence of over 90% in 2002 when four fungicide applications were made, but only reached a maximum of 60% in 2003 when 6 applications were made (Table 1). Pepper spot was generally most severe on early-maturing entries (maturity group = 2). Levels of pepper spot on the maturity group 2 entries generally did not differ from the susceptible cultivar Tamspan 90. The runner-type reference cultivars Okrun and Tamrun 96 had lower levels of pepper spot than Tamspan 90 each year. Entries with the lowest level of pepper spot included late maturing (maturity group 4) entries 103, 158, and 464. Mid-maturing entries (maturity group 3) had both resistant (208, 461, 128, 804, 92, 766) and susceptible (176, 377, 379, 380, 391) phenotypes.
Web blotch appeared late in the season in 2003 and developed to moderate levels on some of the entries. There was considerable variation in genotype reactions to web blotch in 2003 (Table 1). The reference cultivar Tamspan 90 is considered susceptible to web blotch and had an incidence of almost 25%. The runner-type reference cultivars and entries 464, 461, 128, 92, 766, 398, 103, 399, 820, 60, and 785 did not have web blotch symptoms. These and other entries with a low incidence of web blotch were from various maturity and growth habit groups. Defoliation levels from foliar disease averaged 14% in 2002 and 8% in 2003 (data not shown). Tamspan 90, the most susceptible reference cultivar to foliar diseases, had only 15% defoliation in 2002, and less than 10% in 2003. In 2002, entries 426, 437, 60, 570, and 73 had defoliation levels ranging from 35 to 55% and were the only entries that differed from Tamspan 90. In 2003, only entries 129 (26% defoliation) and 73 (21% defoliation) had defoliation levels that differed from Tamspan 90 (8% defoliation).
The average yield in 2003 was 23% higher than in 2002. Yields also were more variable in 2002 (c.v. = 23.4%) compared to 2003 (c.v. = 12%). Yields of Tamrun 96 were highest each year, and significantly greater than the Sclerotinia blight-susceptible cultivar Okrun (Table 1). Tamspan 90 yielded less than Tamrun 96 in 2002, but not in 2003. All core entries yielded less than Tamrun 96 in 2002 and 2003 except for entry 799. However, entry 799 was among the most susceptible entries to Sclerotinia blight in 2003, not differing in disease incidence from Okrun or Tamrun 96. Of the entries that had less than 10% incidence of Sclerotinia blight each year; 361 and 377 had yields that did not differ from Tamspan 90 in 2002; 103, 273, and 460 had yields that did not differ from Tamspan 90 in 2003; and 374, 380, and 562 had yields that did not differ from Tamspan 90 in 2002 and 2003.
Discussion
A wide range of reactions to Sclerotinia blight was observed in evaluation of the peanut core collection in naturally infested fields. Over 10% of the entries had no Sclerotinia blight in 2001 and nearly 30% of the entries had levels of disease from 0 to 10% that would be similar to the resistant reference cultivar Tamspan 90. Conversely, only about 21% of the entries had a disease incidence of 50% or greater that would be similar to runner cultivars such as Okrun and are considered susceptible to Sclerotinia blight. The other half of the collection was intermediate in reaction. Making initial selections from the planting of the entire core collection in a non-replicated trial in 2001 was generally successful. Most of the entries selected in 2001 for further evaluation (42 of 77) had levels of Sclerotinia blight that did not differ from the resistant check cultivar Tamspan 90 in two years of testing in replicated trials. However, there were 15 entries selected in 2001 that had moderate to high incidence of Sclerotinia blight in 2002 and were dropped from further evaluation. In 2003 when disease incidence was high, another nine entries had moderate to high levels of Sclerotinia blight that did not differ from Okrun and/or Tamrun 96. These entries may have been escapes or may have been strongly influenced by environmental conditions given the significant effects of year and year x entry interaction observed for incidence of Sclerotinia blight in 2002 and 2003. In 2002, plant growth was noticeably less vigorous than in 2001 and 2003 as vines of the runner-type reference cultivars Okrun and Tamrun OL02 generally did not overlap between adjacent rows. Environmental effects on plant growth habit may have produced a variable expression of architectural resistance in these entries. Evaluation of all selected entries in an assay to test for physiologic resistance may be beneficial in characterizing the mechanism(s) of resistance. There were over 40 entries that did not have Sclerotinia blight in 2001, but were not kept for further evaluation because their growth habit and vigor was not desirable. These may represent additional sources of resistance not yet thoroughly evaluated.
To our knowledge, this is the first comprehensive screening of plant introductions (PIs) for resistance to Sclerotinia blight. Others have reported disease reactions of a few individual entries in the field. Coffelt (6) found PI 362130 and PI 371961 to be susceptible (>45% disease) while PI 371521 was resistant (11% disease) and similar in response to Sclerotinia blight to the spanish cultivar Chico. PI 371521 is in the core collection as entry 255, but it was one of the entries not received for evaluation in this study. Porter et al. reported that PI 262090, PI 262000, and PI 275751 had good resistance to Sclerotinia blight (5 to 12% disease) while PI 343392 (45 to100% disease) was susceptible (23). PI 261924 was evaluated as core entry 204 in 2001 and had a similar low incidence of Sclerotinia blight (<5%) in our study, but it was not retained for further evaluation. The other plant introductions are not in the core collection. Chenault et al. (5) reported on PI entries from Peru that were either susceptible at >50% incidence of Sclerotinia blight (PI 497598, PI 497669, PI 501273, and PI 502039) or resistant at 10 to 17% disease incidence (PI 476016, PI 501983, PI 501996, PI502009, and PI 502154). None of these entries were in the core collection.
The best levels of partial resistance to Sclerotinia blight adapted to the southwestern U.S. have come from upright spanish (Toalson, Comet) and valencia (UF 734002) germplasm found in the genetic backgrounds of TXAG4 and TXAG5 breeding lines (31), Tamspan 90, and Southwest Runner. However, aside from Tamrun 98, whose resistance was derived from TXAG-5 but was never widely grown because of its low yield potential, crosses with this material have not led to the development of new runner cultivars with partial resistance to Sclerotinia blight. The moderate levels of partial resistance to Sclerotinia blight in the runner type Tamrun 96 and its derivatives Tamrun OL01 and Tamrun OL02 that have a high oleic to linoleic acid oil ratio used another source of disease resistance (PI 475871). Therefore an emphasis of this study was to identify sources of resistance with a prostrate growth habit more amenable to the development of high-yielding runner cultivars. Unfortunately, most of the entries with good resistance had upright growth habits (growth habit = 5) compared to the runner-type reference cultivars and incidence of Sclerotinia blight was negatively correlated with growth habit score. However, most (69%) of the entries in the collection that were received had upright growth habits of 5 or 6. Only 11 of the 62 entries retained for evaluation in both 2002 and 2003 had prostrate (growth habit = 3) or intermediate (growth habit = 4) growth habits and most of these were among the most susceptible of the core entries. However, entries 208 and 582 (growth habit = 3), and 273 (growth habit = 4) were among the resistant entries and were exceptions to the apparent overall trend for architectural resistance. Entry 208 from Bolivia is a spreading bunch type that is susceptible to northern and peanut root-knot nematodes and tolerant to TSWV (USDA/ARS Germplasm Resources Information Network). Entry 582 is a bunch type from Argentina and entry 273 is a spreading type from Malawi that is susceptible to northern and peanut root knot nematodes.
Pepper spot and web blotch are generally considered minor foliar diseases of peanut, but pepper spot occurred consistently all three years of the study and web blotch was present in 2003. In Oklahoma, pepper spot is most commonly observed on certain cultivars and in drier than average years when early leaf spot and web blotch are less severe. Spanish cultivars and Southwest Runner have typically been susceptible to pepper spot while runner types are generally resistant. Tamspan 90 is considered susceptible to both pepper spot and web blotch (8). Fungicide programs have generally not provided good control of pepper spot, but the disease does not cause high levels of defoliation and thus is not thought to be yield limiting. Nevertheless, susceptibility to pepper spot is not a desirable trait and information on entry reaction might be beneficial for breeding programs. In 2001, pepper spot was negatively correlated (P < 0.01) with maturity group (P = −0.55) and positively correlated with growth habit (P = 0.01). Thus, early-maturing entries with upright growth habits are generally most susceptible. Entry reaction to web blotch followed similar trends. Defoliation from foliar disease(s) can influence canopy microclimate by effectively opening the canopy resulting in more light penetration and air movement, which are typically less favorable for Sclerotinia blight development. Canopy modification by foliar disease can severely limit Sclerotinia blight development and has been most apparent in fungicide trials where early leaf spot is allowed to cause severe defoliation in untreated check plots. However, mean defoliation in 2002 was only 14% and exceeded 25% for only 12 of the entries. In 2003 mean defoliation was only 8% and only exceeded 20% for 2 entries. Thus, the effects of foliar diseases on Sclerotinia blight development during the course of the study were considered minimal.
The criteria of resistance to Sclerotinia blight, yield similar to Tamspan 90, growth habit, and/or pepper spot and web blotch reaction were used to select entries for further evaluation. Entries 208, 128, 804, 582, and 273 combined resistances to Sclerotinia blight, pepper spot and web blotch; and less than erect growth habits. Entry 103 had good Sclerotinia blight resistance and yield, but an upright growth habit. Entry 92 had an upright growth habit and low yield, but good Sclerotinia blight resistance. Entries 92 and 103 had upright growth habits but were among the best entries for resistance to web blotch and pepper spot. Entries 426, 184, and 562 were upright and susceptible to susceptible to pepper spot, but had some resistance to web blotch and had the best resistance to Sclerotinia blight. Entry 562 also had good yields. These entries appear to be useful sources of resistance to Sclerotinia blight for breeding programs and for increasing the probability of finding additional sources of resistance in clusters of additional germplasm identified within the entire USDA collection.
Chenault et al. (5) identified a molecular marker for identifying resistance to Sclerotinia blight in peanut. Using a pair of simple sequence repeat primers, 145 and 100 base pair (bp) bands were produced and associated with resistance and susceptibility, respectively. Various genotypes of either band alone, or combinations of both bands were associated with resistant and susceptible phenotypes among a wide range of runner, spanish, and valencia entries. However, the marker was not present in several virginia entries with good resistance. Core entries 103, 128, 208, and 273 that were selected from this study based on their resistance; and the reference cultivars Okrun, Tamrun 96, and Tamspan 90 were tested using polymerase chain reaction in the study by Chenault et al. (5). Core entries 103, 128, 208, and 273; and Tamrun 96 each only had the 145 bp band for resistance. Tamspan 90 had both bands with the 145 bp band for resistance dominating and Okrun only had the 100 bp band for susceptibility. None of the susceptible core entries from this study were tested for the marker. The 145 pb band for resistance was frequently found in prostrate plant types, possibly indicating the involvement of physiologic resistance. Inheritance of resistance to Sclerotinia blight using TX-AG5 is not clearly understood but appears to be controlled by at least two loci (37) with low to moderate heritability estimates ranging from 14 to 47% (7,37). The use of molecular marker technology during a recurrent backcrossing program may expedite the development of cultivars with improved resistance to Sclerotinia blight using the sources of resistance identified in this study and those yet to be identified in the entire peanut collection.
Literature Cited
Akem C. N. , Melouk H. A. , and Smith O. D. 1992 Field evaluation of peanut genotypes for resistance to Sclerotinia blight. Crop Prot 11 : 345 – 348 .
Anderson W. F. , Holbrook C. C. , and Culbreath A. K. 1996 Screening the peanut core collection for resistance to tomato spotted wilt virus. Peanut Sci 23 : 57 – 61 .
Banks D. J. , Kirby J. S. , and Sholar J. R. 1989 Registration of ‘Okrun’ peanut. Crop Sci 29 : 1574 .
Chappell G. F. , Shew B. B. , Ferguson J. M. , and Beute M. K. 1995 Mechanisms of resistance to Sclerotinia minor in selected peanut genotypes. Crop Sci 35 : 692 – 696 .
Chenault K. D. , Maas A. L. , Damicone J. P. , Payton M. E. , and Melouk H. A. 2009 Discovery and characterization of a molecular marker for Sclerotinia minor (Jagger) resistance in peanut. Euphytica 166 : 357 – 365 .
Coffelt T. A. and Porter D. M. 1982 Screening peanuts for resistance to Sclerotinia blight. Plant Dis 66 : 385 – 387 .
Cruickshank A. W. , Cooper M. , and Ryley M. J. 2002 Peanut resistance to Sclerotinia minor and S. sclerotiorum. Aust. J. Agric. Res 53 : 1105 – 1110 .
Damicone J. P. and Jackson K. E. 1996 Disease and yield responses to fungicides among peanut cultivars differing in reaction to Sclerotinia blight. Peanut Sci 23 : 81 – 85 .
Damicone J. P. and Jackson K. E. 2001 Effects of application method and rate on control of Sclerotinia blight with iprodione and fluazinam. Peanut Sci 28 : 28 – 33 .
Damicone J. P. , Jackson K. E. , and Dashiell K. E. 2004 Responses of peanut cultivars to fluazinam and boscalid for control of Sclerotinia blight. Proc. Amer. Peanut Res. and Educ. Soc 36 : 50 – 51 (abstr.).
Damicone J. P. , Jackson K. E. , Dashiell K. E. , Melouk H. A. , and Holbrook C. C. 2003 Reaction of the peanut core collection to Sclerotinia blight and pepper spot. Proc. Amer. Peanut Res. and Educ. Soc 35 : 55 (abstr.) .
Frankel O. H.
1984
Genetic perspectives of germplasm conservation.
161
–
170
In
Arber W.
,
Llimensee K.
,
Peacock W. J.
, and
Starlinger P.
(eds.),
Genetic Manipulations: Impact on man and Society
Cambridge Univ. Press
.
Franke M. D. , Brenneman T. B. , and Holbrook C. C. 1999 Identification of resistance to Rhizoctonia limb rot in a core collection of peanut germplasm. Plant Dis 83 : 944 – 948 .
Godsey C. , Damicone J. , Mulder P. , Medlin C. , Kizer M. , Noyes R. , and Zhang H. 2008 Peanut production guide for Oklahoma. Okla. Coop. Ext. Serv. Cir. E-808. 87 .
Goldman J. J. , Smith O. D. , Simpson C. E. , and Melouk H. A. 1995 Progress in breeding Sclerotinia blight-resistant runner type peanut. Peanut Sci 22 : 109 – 113 .
Holbrook C. C. 2001 Status of the Arachis germplasm collection in the United States. Peanut Sci 28 : 84 – 89 .
Holbrook C. C. and Anderson W. F. 1995 Evaluation of a core collection to identify resistance to late leafspot in peanut. Crop Sci 35 : 1700 – 1702 .
Holbrook C. C. , Anderson W. F. , and Pitman R. N. 1993 Selection of a core collection from the U.S. germplasm collection of peanut. Crop Sci 33 : 859 – 861 .
Holbrook C. C. , Timper P. , and Xue H. Q. 2000 Evaluation of the core collection approach for identifying resistance to Meloidogyne arenaria in peanut. Crop. Sci 40 : 1172 – 1175 .
Hollowell J. E. , Shew B. B. , and Isleib T. G. 2003 Evaluating isolate aggressiveness and host resistance from peanut leaflet inoculations with Sclerotinia minor. Plant Dis 87 : 402 – 406 .
Hollowell J. E. , Isleib T. G. , Tallury S. P. , Copeland S. C. , and Shew B. B. 2008 Screening of virginia-type peanut breeding lines for resistance to Cylindrocladium black rot and Sclerotinia blight in the greenhouse. Peanut Sci 35 : 18 – 24 .
Kirby J. S. , Melouk H. A. , Stevens T. E. , Banks D. J. , Sholar J. R. , Damicone J. P. , and Jackson K. E. 1998 Registration of ‘Southwest Runner’ peanut. Crop Sci 38 : 545 – 546 .
Porter D. M. and Beute M. K. 1974 Sclerotinia blight of peanuts. Phytopathology 64 : 263 – 264 .
Porter D. M. , Beute M. K. , and Wynne J. C. 1975 Resistance of peanut germplasm to Sclerotinia sclerotiorum. Peanut Sci 2 : 78 – 80 .
Porter D. M.
and
Melouk H. M.
1997
Sclerotinia blight.
34
–
35
in:
Compendium of Peanut diseases, 2nd. ed
Kokalis-Burelle N.
,
Porter D. M.
,
Rodriguez-Kabana R.
,
Smith D. H.
, and
Subrahmanyam P.
(eds.),
American Phytopathological Society Press
St. Paul, MN
.
Phipps P. M. 1995 An assessment of environmental conditions preceding outbreaks of Scletrotinia blight of peanut in Virginia. Peanut Sci 22 : 90 – 93 .
Saxton A. M. 1998 A macro for converting mean separation output to letter groupings in Proc Mixed In Proc. 23rd SAS Users Group Intl., SAS Institute Cary, NC 1243 – 1246, Nashville, TN, March 22–25.
Simpson C. E. , Baring M. R. , Shubert A. M. , Melouk H. A. , Black M. C. , Lopez Y. , and Keim K. A. 2003 Registration of ‘Tamrun OL01’ peanut. Crop Sci 46 : 2298 .
Simpson C. E. , Baring M. R. , Shubert A. M. , Black M. C. , Melouk H. A. , and Lopez Y. 2006 Registration of ‘Tamrun OL02’ peanut. Crop Sci 46 : 1813 – 1814 .
Simpson C. E. , Smith O. D. , and Melouk H. A. 2000 Registration of ‘Tamrun 98’ peanut. Crop Sci 40 : 859 .
Smith O. D. , Aguirre S. M. , Boswell T. E. , Grichar W. J. , Melouk H. A. , and Simpson C. E. 1990 Registration of TXAG-4 and TXAG-5 peanut germplasms. Crop Sci 30 : 429 .
Smith O. D. , Simpson C. E. , Grichar W. J. , and Melouk H. A. 1991 Registration of ‘Tamspan 90’ peanut. Crop Sci 31 : 1711 .
Smith O. D. , Simpson C. E. , Black M. C. , and Besler B. A. 1998 Registration of ‘Tamrun 96’ peanut. Crop Sci 38 : 1403 .
Smith D. L. , Garrison M. C. , Hollowell J. E. , Isleib T. G. , and Shew B. B. 2008 Evaluation of application timing and efficacy of the fungicides fluazinam and boscalid for control of Sclerotinia blight of peanut. Crop Protection 27 : 823 – 833 .
Smith D. L. , Hollowell J. E. , Isleib T. G. , and Shew B. B. 2006 Analysis of factors that influence the epidemiology of Sclerotinia minor on peanut. Plant Dis 90 : 1425 – 1432 .
Wadsworth D. F. 1979 Sclerotinia blight of peanuts in Oklahoma and the occurrence of the sexual stage of the pathogen. Peanut Sci 6 : 77 – 79 .
Wildman L. G. , Smith O. D. , Simpson C. E. , and Taber R. A. 1992 Inheritance of resistance to Sclerotinia minor in selected spanish peanut crosses. Peanut Sci 19 : 31 – 34 .
Notes
Approved for publication by the Director of the Oklahoma Agricultural Experiment Station. This research was funded in part by the Oklahoma Agricultural Experiment Station, the Oklahoma Peanut Commission, and the National Peanut Board.
Author Affiliations
1 Professor and Assistant Professor, Department of Entomology and Plant Pathology, Oklahoma State University, Stillwater, OK 74078-3033.
2 Research Leader and Research Geneticist, USDA/ARS, Tifton, GA 31793.
3 Research Plant Pathologist and Research Geneticist, respectively, USDA/ARS, Stillwater, OK 74075-2714.