

Introduction
Improved productivity of available agricultural land resources is a global concern. Moreover, farmers seek options to sustain their livelihood in the face of increased input costs (Moss et al., 2017). Interest has increased in harvesting market type peanuts for forage (Foster et al., 2012) because it can yield as much as 6 Mg/ha (Santos and Sutton, 1982; Sorensen et al., 2009) of high quality forage, as indicated by a high CP concentration (Foster et al., 2012; Prine, 1964). Still, because market type peanuts are more valuable as human food than livestock feed and establishment costs are high, if any forage is harvested, it is usually harvested as a stover baled after the pods are threshed (Santos and Sutton, 1982; Sorensen et al, 2009; Yang, 2005). Except for CP, the nutritive value of peanut stover is similar to full-bloom alfalfa (Medicago sativa L.) (Yang, 2005) with 80 g/kg CP, 350 g/kg acid detergent fiber (ADF), and 430 g/kg neutral detergent fiber (NDF) (Packard et al., 2007). Peanut forage harvested prior to pod maturity (at R2 vs. R8, Boote, 1982) had greater nutritive value (i.e. 177 g/kg CP and 270 g/kg ADF; Foster et al., 2012); but pod yield, which is more valuable than forage, can be reduced (Prine, 1964; Santos and Sutton, 1982). Santos and Sutton (1982) found that the timing of forage harvest determined the effect, if any, on pod yield of Virginia bunch market types. Sorensen et al. (2009) also concluded that early season (8.5 WAP) harvest of forage from a runner market type was not economically feasible but that a later season (13-17 WAP) single harvest was feasible and that, while the single harvest produced equal yield to multiple forage harvests, it also minimized the effect on pod yield at 18-22 WAP compared with multiple harvests.
Valencia market type landraces and improved cultivars are used on every continent except Antarctica (Dwivedi et al., 2008). In the USA, Valencia market types are predominantly grown in eastern New Mexico and west Texas (the Southern High Plains) for the in-shell market with average annual pod yields of 3750 kg/ha when harvested 22 WAP (Dwivedi et al., 2008). Valencia market type producers may gain added value by harvesting peanut forage before digging pods to be sold as high quality hay as opposed to baling lower quality stover after pods are dug, sun-cured, and threshed; however, information is limited on peanut forage and pod production (Sorensen et al., 2009) and previously non-existent for the Valencia market type. The objective of this study was to evaluate the effects of selected single harvest timing in relation to pod harvest maturity on the forage yield and nutritive value and pod yield of Valencia market type peanut.
Materials and Methods
Locations, Tillage, Cultivar, and Plot Size
Research was conducted on two separate producer fields near Brownfield, TX, USA (Sites 1 and 2) in 2012 and 2013. The experimental design was a randomized complete block with four replications within each location × year combination. Valencia peanut cultivar Valencia C, release by New Mexico State University (Hsi, 1980) was planted (20 seeds/m) in mid-May each year into conventionally tilled seedbeds that had been formed into 1-m beds. Plots were 3.67 m × 1 bed. Management of the experiment was under producer control. Weeds, insects, and diseases were controlled or prevented as needed with labeled products each year (Sorensen et al., 2009); fertilizers were applied based on soil test recommendations, and sprinkler irrigation using groundwater was applied as needed to prevent moisture stress. Digging took place 22 WAP during the third week of October when peanut was at harvest maturity (R8) based on the hull scrape method (Boote, 1982; Sorensen et al., 2009; Williams and Drexler, 1981). Weather data from the nearest available station are shown for each year in Table 1.

Weather dataa during 2012 and 2013 and the long-term averages near Brownfield, TX, USA.
Experimental Treatments and Data Collection
Treatments were forage harvests taken during the R7.5 to R8 pod maturity stages (Boote, 1982), 18, 20, 21, and 22 WAP. Standing forage above 5 cm on bed tops was cut with a sickle-bar mower and weighed in the field. A sample from each plot was weighed, dried at 65 C for 48 h, and reweighed to determine dry matter (DM) concentration and to convert field weights to DM yield. Forage from the 22 WAP treatment was intact at the time of digging and left at the soil surface after digging to cure in the field until threshing in early November (Burkey et al., 2007; Sorensen et al., 2009) at which time it was collected from the thresher, weighed, and subsampled for DM as previously described. Forage samples from both sites in 2012 were ground to pass through a 1-mm screen and analyzed by near infrared reflectance spectroscopy (NIRS) to estimate forage nutritive value components (Lauriault and Kirksey, 2004; Marsalis et al., 2008) using an equation developed by Ward Laboratories (Kearney, NE, USA) from a subset of samples from this study. Ash estimates were used to convert DM yield to OM yield. After threshing, pods were air-dried and weighed.
Statistical Analysis
Forage organic matter and pod yield data were analyzed with the Mixed procedure of SAS (SAS Inst., 2011). Tested effects included year, site, WAP, and all possible interactions. Nutritive value data [CP, ADF, NDF, and NDF digestibility (NDFD)] also were analyzed with the Mixed procedure of SAS (SAS Inst., 2010). Tested effects included WAP, site, and their interaction. Rep × year × site and residual mean squares were considered random (Littell et al., 2002). All differences reported are significant at P ≤ 0.05. When a main effect or interaction was significant, protected (P ≤ 0.05) least significant differences were used to determine where differences occurred among treatment means using the PDMIX800 SAS macro (Arnold M. Saxton, University of Tennessee, Knoxville, 2000).
Results and Discussion
Forage organic matter yield
Main effects of year, site, and WAP were significant for forage organic matter yields, which declined across WAP (Table 2). Pimratch et al. (2008) found no differences between years in biomass production. Forage OM yields observed in this study were consistent to or slightly lesser than those reported by Foster et al. (2012) who used the same stubble height for runner type peanut, but did not convert to OM yield. Forage OM yields in the present study were considerably greater than those reported by Sorensen et al. (2009) likely because they took earlier harvests, which limited biomass accumulation, and because they used a much taller stubble height (20 cm).
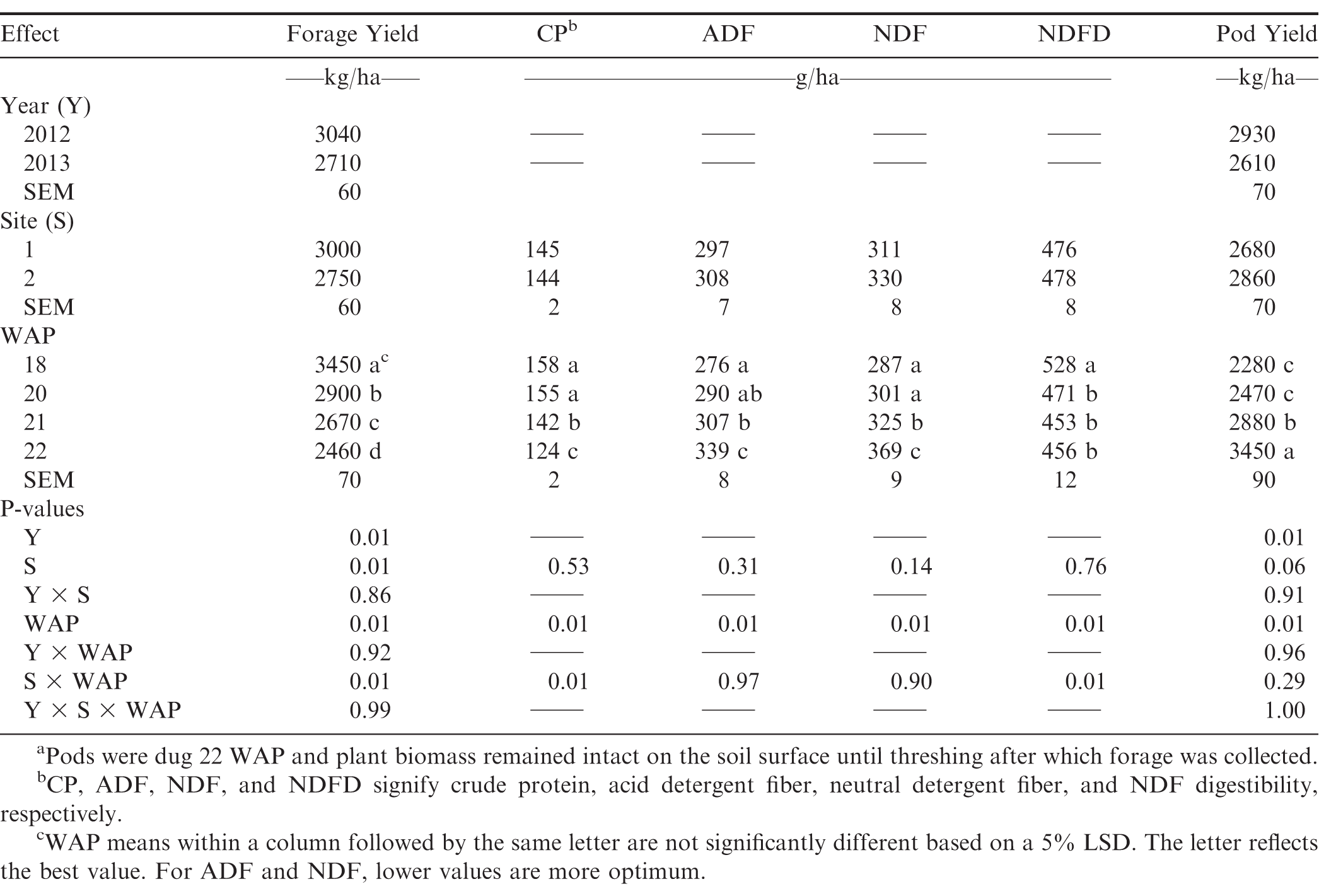
Forage organic matter yield and forage nutritive value and pod yield of Valencia peanut when forage was harvested 18, 20, 21, and 22a weeks after planting (WAP) at 2 sites near Brownfield, Texas each in 2012 and 2013. Data are the LSMeans of 4 replicates in each site-year.
The significant site × WAP interaction, shown in Table 3, was due to a difference in yield at 18 WAP and a difference in the rate of decline in yield from 18 to 22 WAP, likely due to differences in producer management for irrigation, fertility, pest control, and harvesting. Sorensen et al. (2009) and Santos and Sutton (1982) also observed an increase in leaf and stem yield through 17 and 18 WAP, respectively. Santos and Sutton (1982) also reported a decline in forage yield of Virginia bunch market type from 18 to 22 WAP, at which time pod harvest maturity (R8; Boote, 1982) had occurred, making that a similar timeframe of harvests to the present study as well as to Sorensen et al. (2009) for the 13 to 17 WAP treatments. Foster et al., (2012) measured greater yield when peanut forage was harvested at R2 compared to R8, attributing the difference to taller plants and stem growth at R2 that senesced by R8 (Boote, 1982). Santos and Sutton (1982) attributed the forage yield decline to a combination of leaf senescence and falling, and repartition of assimilates from vegetative top growth to reproductive functions, which may have been the case in the present study. Santos and Sutton (1982) stated that a reduction in leaf: stem ratio was coincidental to the leaf drop; however, Sorensen et al. (2009) measured a reduction in leaf: stem ratio of single-cut peanut forage as the harvest was delayed and attributed it to stem and internode elongation. Boote et al. (1980) found that peanut canopies can sustain pod yield with some defoliation; but, that lack of photosynthetic production due to defoliation could lead to a reduction in stem yield as assimilates will be reallocated to seed filling. Sorensen et al. (2009) suggested that, instead of allowing the leaves to drop, they could be harvested along with the stems before pod harvest for higher quality hay.
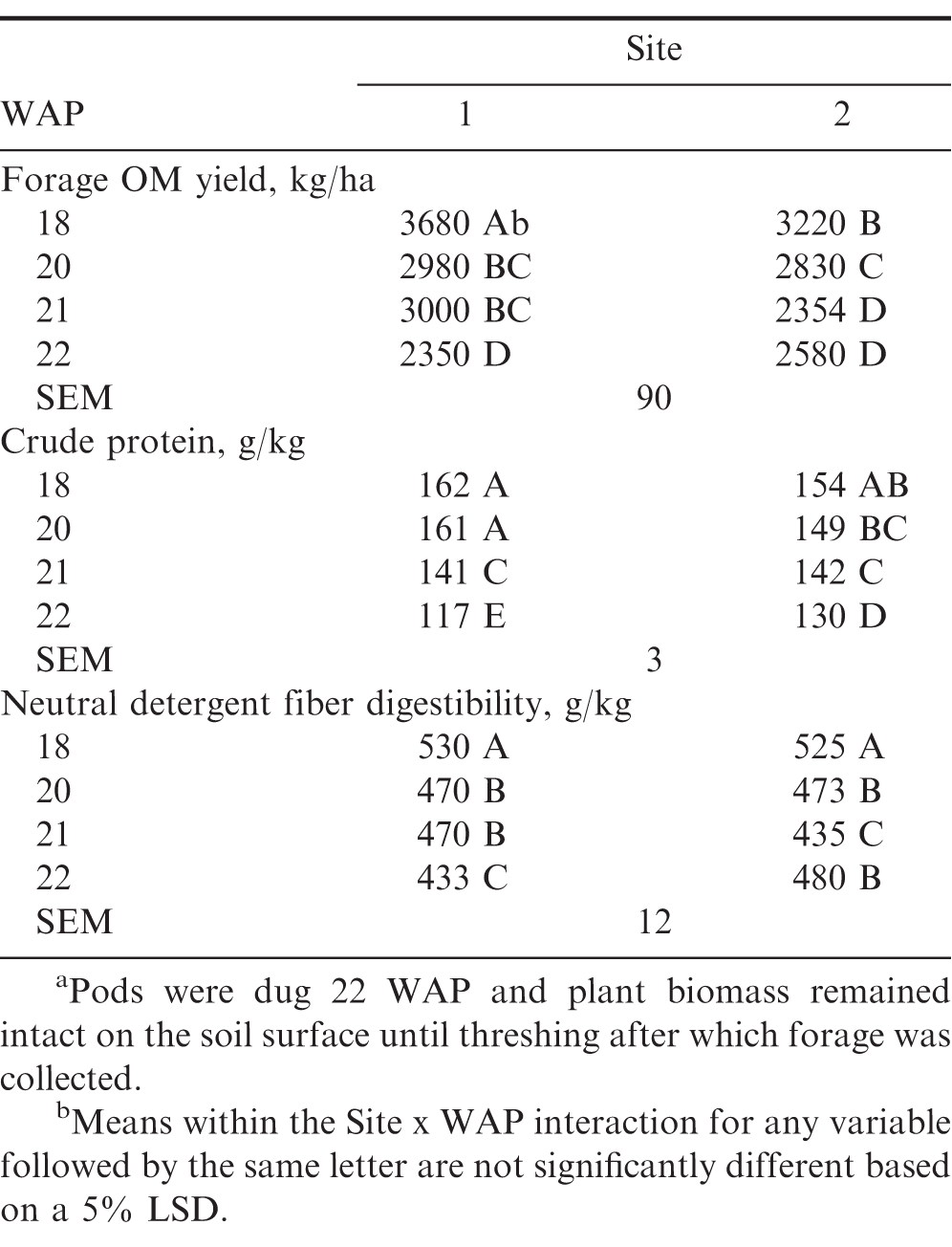
Forage organic matter (OM) yield, crude protein, and neutral detergent fiber digestibility of Valencia peanut when forage was harvested 18, 20, 21, and 22a weeks after planting (WAP) at 2 sites in a single year near Brownfield, Texas. Data are the LSmeans of 4 replicates for each WAP forage harvest treatment at each site.
Sorensen et al. (2009) observed greater yield of forage harvested after digging, compared to pre-digging forage harvest treatments, suggesting that forage harvested after digging, curing, and threshing included pods that bypassed the thresher in addition to root mass and soil. Organic matter yields are reported for the present study to avoid the inclusion of soil.
Forage nutritive value components
In the present study CP declined as time progressed from 21 to 22 WAP (Table 2). Crude protein concentration of the peanut forage in this study estimated by NIRS (Table 2) was consistent to that measured by conventional wet chemistry methods by Foster et al. (2012) for peanut harvested at R8 (22 WAP in the present study, 120 g/kg CP) and perennial peanut (A. glabrata L.) (156 g/kg CP) measured by Prine (1964) in late summer/early autumn, which would correspond to 18 and 20 WAP in the present study. As with OM yield, the significant site × WAP interaction was due to difference in the rate of decline of CP between sites (Table 3), again, likely due to differences in producer management for irrigation, fertility, pest control, and harvesting.
Both ADF and NDF increased as pod digging approached (Table 2), although, as with CP, the decline did not occur until 20 WAP. Acid detergent fiber measured at 21 and 22 WAP in this study was similar or slightly greater than that measured by Foster et al. (2012) (309 g/kg ADF) at R8 (Boote, 1982). Neutral detergent fiber in this study measured at R8 (Boote, 1982) was lower than that measured by Foster et al. (2012) (443 g/kg NDF). These nutritive value components (CP, ADF, and NDF) were all more optimum than those reported by Packard et al. (2007) for sun-cured peanut stover hay as 80, 350, and 430 g/kg CP, ADF, and NDF, respectively.
Neutral detergent fiber digestibility declined after 18 WAP (Table 2). There was a difference in the rate of decline of NDFD between sites (Table 3) likely due to differences in producer management and is not considered to be of biological importance. Foster et al. (2012) did not report NDFD; however, they did report that while fiber fractions (including ADF and NDF) increased from R2 to R8 (Boote, 1982), there was no effect of harvest maturity on in vitro true digestibility (IVTD). They (Foster et al., 2012) attributed the lack of difference in IVTD at various forage harvest maturities to relatively small differences in ADF and lignin and no difference in water soluble carbohydrates. Each of these changes in nutritive value can be attributable to leaf senescence and drop (Santos and Sultan, 1982), stem elongation (Sorensen et al., 2009), and reallocation of assimilates from vegetative to reproductive components (Santos and Sultan, 1982).
Pod yield
Pod yield in the present study at 22 WAP (Table 2) was less than the long-term average regional yields for Valencia peanuts reported by Dwivedi et al. (2008). Pod yield differed across years by 11%, which is much less than the 25% observed by Burkey et al. (2007) in growth chamber studies and the nearly 40% reported by Sorensen et al. (2009). Sorensen et al. (2009) conducted field studies in which the yield difference across years was attributed to drought, which was not likely a factor in the present study due to irrigation practices typical of the area that prevent moisture stress. While Burkey et al. (2007) and Sorensen et al. (2009) both observed a sizable difference between years in pod yield, Booker et al. (2007) in a companion study combined years for aboveground biomass data, possibly due to a lack of difference between years. There also was a trend (P < 0.06) toward a difference in pod yield between sites that was a reversal of the difference in forage OM yield (Table 2). It is possible that one producer's management supported forage production over pod production while the other was more favorable to pod production.
Pod yield (Table 2) increased with delayed forage harvest after 20 WAP, but it was considerably less than those measured in Georgia by Sorensen et al. (2009) and by Moss et al. (2017), both of whom used a runner type. Foster et al. (2012) did not report any effects of harvest at R2 or R8 on pod yield. All forage harvest treatments imposed by Sorensen et al. (2009) prior to pod digging reduced pod yield compared with forage harvest after digging. Nonetheless, there was a nonsignificant increase in pod yield as harvest was delayed before digging in that study that was somewhat consistent to the increase in yield observed in the present study, albeit at lesser yields (Table 2). Pod yield was reduced by 34% in the present study when forage was harvested at 18 WAP (calculated from Table 2) compared with 3% observed by Sorensen et al. (2009) who did not take forage harvests later than 17 WAP, but did note increasing pod yield with single harvests up to that time. Sorensen et al. (2009) used a 20-cm cutting height. The 5-cm height used in the present study may have severed pegs preventing continued allocation of assimilates for increased pod-filling.
Santos and Sutton (1982) measured reduced pod yields by harvesting forage between pod formation and pod-filling. Harvest at flowering did not reduce yields and forage harvests were not taken later than 18 WAP (R6; Boote, 1982) in that study (Santos and Sutton, 1982) as was done in the present study (Table 2). Boote et al. (1980) concluded that defoliation during seed-filling was most detrimental and that earlier defoliation permits regrowth before the critical period. Based on the present study, it may be that forage harvests during the approximately 30-d pod maturation process (R7 to R8; Boote, 1982), which was later than 18 WAP (Santos and Sutton, 1982) and 17 WAP (Sorensen et al., 2009), had a detrimental effect on pod yield at R8 (pod harvest maturity; Boote, 1982). In fact, at the 2013 prices of $0.73/kg and $0.084/kg for New Mexico in-shell peanut and hay excluding alfalfa (https://quickstats.nass.usda.gov/), gross receipts for the 18, 20, 21, and 22 WAP forage harvest treatments totaled $1963, 2059, 2306, and 2745/ha.
Conclusions
Harvesting Valencia peanut for hay during the pod-filling stage (18 WAP or after) reduced forage yield and nutritive value compared to harvesting at 18 WAP; however, pod yield was decreased by earlier compared to later forage harvests or harvesting hay after digging and threshing. Others, using different peanut types in different environments, have shown that earlier harvests also reduced forage and pod yield compared to harvesting hay after digging and threshing. Consequently, forage harvest of Valencia market type peanut within five weeks of pod harvest is not likely a viable option for producers to increase revenue. Future studies may reveal that earlier forage harvests or leaving a taller stubble may be feasible to not reduce pod yields of Valencia market type peanuts.
Acknowledgements
We gratefully acknowledge the field assistance of Calvin Henson, Gilbert Lucero, Martin Mead, Larry Perkins, Kenneth Phipps, and Aaron Scott; office assistance from Patty Cooksey and Valerie Pipkin; and our coworkers at the NMSU Library Document Delivery Service. Salaries and research support were provided by state and federal funds appropriated to the New Mexico Agricultural Experiment Station.
Literature Cited
F.L., Booker, K.O Burkey, W.A Pursely, and A.S Heagle (2007). Elevated carbon dioxide and ozone effects on peanut: I. Gas-exchange, biomass, and leaf chemistry. Crop Sci 47: 1475- 1487.
K.J Boote, (1982). Growth stages of peanut (Arachis hypogaea L.). Peanut Sci 9: 35- 40.
K.J., Boote, J.W Jones, G.H Smerage, C.S Barfield, and R.D Berger (1980). Photosynthesis of peanut canopies as affected by leafspot and artificial defoliation. Agron. J 72: 247- 252.
K.O., Burkey, F.L Booker, W.A Pursely, and A.S Heagle (2007). Elevated carbon dioxide and ozone effects on peanut: II. Seed yield and quality. Crop Sci 47: 1488- 1497.
S.L., Dwivedi, N Puppala, H. D Upadhyaya, N Manivannan, and S Singh (2008). Developing a core collection of peanut specific to Valencia market type. Crop Sci 48: 625- 632.
J.L., Foster, G.C Lamb, B.L. tillman, J.J Marois, D.L Wright, and M.K Maddox (2012). In sacco degradation kinetics of fresh and field-cured peanut (Arachis hypogaea L.) forage harvested at different maturities. Anim. Feed Sci. Tech 171: 52- 59.
D.C. Hsi, (1980). Regisration of New Mexico Valencia C. Crop Sci 20: 113- 114.
L.M., Lauriault, and R.E Kirksey (2004). Yield and nutritive value of irrigated winter cereal forage grass-legume intercrops in the southern high plains, USA. Agron. J 96: 352- 358.
R.C., Littell, W.W Stroup, and R.J Freund (2002). SAS for linear models, 4th ed. 466 pp. SAS Inst. Inc. Cary, NC USA..
Marsalis, M.A., L.M Lauriault, and D.M VanLeeuwan 2008 Selecting small-grain forages for the southern High Plains Online Forage and Grazinglands doi:10.1094/FG-2008-1104-01-RS.
Moss, J.W., R.S Tubbs, T.L Grey, N.B Smith, and J.W Johnson 2017 Assessment of double crop and relay intercropping systems of peanut and soft red winter wheat and residual herbicides. Online Crop, Forage, and Turfgrass Manage DOI: 10.2134/cftm2016.10.0069.
C.E., Packard, J.P Muir, and R.D Wittie (2007). Effects of groundnut stover or bermudagrass hay supplementation to doe kids on winter hardwood range. Small Ruminant Res 67: 1- 6.
S. S Pimratch, Jogloy, N Vorasoot, B Toomsan, A Patanothai, and C.C Holbrook (2008). Relationship between biomass production and nitrogen fixation under drought-stress conditions in peanut genotypes with different levels of drought resistance. J. Agron. Crop Sci 194: 15- 25.
G.M Prine, (1964). Forage possibilities in the genus Arachis. Proc. Soil Crop Sci. Soc. Fla 24: 187- 196.
R.B., Santos, and B.G Sutton (1982). Effect of defoliation on Virginia Bunch peanuts at Camden, N.S.W. Aust. J. Agric. Res 33: 1037- 1048.
SAS Inst 2011 The SAS system for Windows, release 9.3 The SAS Inst., Inc., Cary, NC.
R.B., Sorensen, R.C Nuti, and C.L Butts (2009). Yield and plant growth response of peanut to midseason forage harvest. Agron. J 101: 1198- 1203.
E.J., Williams, and J.S Drexler (1981). A non-destructive method for determining peanut maturity. Peanut Sci 8: 134- 141.
C.-M. J Yang, (2005). Proteolysis, fermentation efficiency, and in vitro ruminal digestion of peanut stover ensiled with raw or heated corn. J. Dairy Sci 88: 2903- 2910.
Notes
- Forage Crop Management Scientist, Plant and Environmental Sciences Dept., New Mexico State University Rex E. Kirksey Agricultural Science Center at Tucumcari, Tucumcari, NM 88401 [^]
- Peanut Breeder, Plant and Environmental Sciences Dept., New Mexico State University Agricultural Science Center at Clovis, 2346 SR 288, Clovis, NM 88101. [^] *Corresponding author Email: npuppala@nmsu.edu
Author Affiliations