

Introduction
Residual herbicide options for peanut include imazapic for postemergence (POST) application, diclosulam for preemergence (PRE) or POST, and flumioxazin for PRE application. Each provides broad-spectrum weed control, but has strengths and weaknesses when used in peanut. With respect to mode of action, flumioxazin (Anderson et al., 1994; Yoshida et al., 1991) is a protoporphyrinogen oxidase inhibitor (PPO) while diclosulam and imazapic inhibit acetolactate synthase (ALS) (Vencill 2002). ALS herbicide resistance (Wise et al., 2009) in Palmer amaranth (Amaranthus palmeri S. Wats.) has increased concerns about proper herbicide stewardship. Concerns about herbicide resistance in other weed species as increased because many herbicides used in the same or in rotational crops, have similar or the same modes of action. Thus, this continuous exposure of the same weeds to the same herbicide modes of action could select for resistance.
Florida beggarweed [(Desmodium tortuosum (Sweet) DC)] emergence occurs throughout the growing season and is a vigorous competitor with peanut if emergence occurs within the first 8 wk of planting (Buchanan et al., 1976; Hauser et al., 1982; 1975; Cardina and Brecke, 1991;). Lack of early season Florida beggarweed control can lead to season-long interference that reduces yield (Grey and Bridges, 2005; Hauser et al., 1975; 1982; Cardina and Brecke, 1991; Wehtje et al., 2000a; 2000b) and hinders crop harvest by interfering with digging and threshing (Buchanan et al., 1982; Wehtje et al., 1999). Florida beggarweed consistently ranks as the most troublesome weed of peanut based on surveys for Alabama, Florida, and Georgia (Buchanan et al., 1982; Elmore, 1989; Dowler, 1992, 1998; Prostko, 2001; Webster and MacDonald, 2001; Webster 2005).
When herbicides are POST applied in peanut, variable Florida beggarweed control can occur. Paraquat provides only early postemergence (EPOST) contact control (Grey et al., 2003), while chlorimuron provides contact and residual activity (Wehtje et al., 2000a). Peanut tolerance to paraquat was first noted in the late 1970s (Wehtje et al., 1986). However, lack of residual activity and the narrow window of application for control of Florida beggarweed reduced the effectiveness of this treatment (Wilcut et al., 1995). Chlorimuron effectively controls Florida beggarweed, but can only be applied 60 d after peanut emergence (Johnson et al., 1992b). This frequently becomes a mid-season salvage treatment (Johnson et al., 1992a). By 60 d after peanut emergence, Florida beggarweed can be taller than the 25 cm in height, which is the maximum specified on the chlorimuron label (Webster et al., 1997; Wehtje et al., 1999). Chlorimuron is still applied to approximately 20% of the Georgia crop (NASS, 2004). Producers have indicated Florida beggarweed control with chlorimuron can be inconsistent. An additional concern is that chlorimuron is an ALS herbicide (Vencil, 2002) with a mode of action similar to other herbicides used in peanut including diclosulam and imazapic.
Studies that have compared the effect of multiple applications of the same mode of action in the same crop for control of Florida beggarweed have not been reported. Thus, research was conducted to evaluate PRE, POST and late POST (LPOST) applications of ALS versus PPO mode of action herbicides for Florida beggarweed establishment, control, population, growth, and competition in peanut.
Materials and Methods
Experiments were conducted in Georgia during 2005 and 2006 at the Ponder Research Farm located near Ty Ty in separate areas of the same field, and in Alabama at the Wiregrass Research and Extension Center near Headland. Soils were Tifton loamy sand (fine-loamy, kaolinitic, thermic Plinthic Kandiudults) at Ty Ty and Dothan fine sandy loam (fine, loamy, siliceous, thermic, Plinthic Paleudult) at Headland. Organic matter ranged from 1.0 to 1.1% with pH from 6.0 to 6.5 at Ty Ty and 1.3% organic matter and pH 6.5 at Headland.
The peanut cultivar Georgia Green (Branch, 1996) was planted in conventionally prepared seedbeds four to five cm deep with seed spaced five to six cm apart at all locations. Planting dates were 10 May 2005 and 2006 at Ty Ty; 17 May 2005 and 15 May 2006 at Headland. Plot size was two rows spaced 91-cm apart by 7.6 m long. Standard cultural practices for peanut production were followed using University of Georgia and Auburn University Extension recommendations. Peanut was irrigated as needed at all locations. Florida beggarweed seed (Adams-Briscoe Seed Co. 325 East 2nd Street, Jackson, GA 30233) were hand sown at approximately 1 kg ha−1 to supplement the naturally occurring population.
Herbicide treatments applied alone were diclosulam (27, 40, and 53 g ha−1) PRE, flumioxazin (53, 80, and 105 g ha−1) PRE, imazapic (36, 52, and 71 g ha−1) POST four wk after peanut emergence, or paraquat plus bentazon (280 + 500 g ha−1) EPOST two wk after peanut emergence. These residual and contact herbicide treatments were applied alone or were followed by chlorimuron at 9 g ai/ha late post emergence (LPOST). Nonionic surfactant at 0.25% (v/v) was included with all EPOST, POST, and LPOST treatments. Herbicides were applied with a CO2-pressurized backpack sprayer calibrated to deliver 140 L/ha at 210 kPa. Each experiment was arranged as a randomized complete block design with a factorial arrangement of treatments replicated four times.
Paraquat plus bentazon EPOST treatments were applied to Florida beggarweed that ranged in size from cotyledon to three leaves and peanut had approximately one to three true leaves. Imazapic POST was applied when Florida beggarweed ranged in size from cotyledon to six leaves with densities ranging from one to five plants m−2 for each location for both years. Peanut had approximately four to seven true leaves when imazapic was POST applied. Chlorimuron LPOST was applied when Florida beggarweed ranged in size from 4 to 10 leaves. Visual estimates of percent Florida beggarweed control were recorded within one wk prior to harvest using a scale of 0 (no control) to 100% (complete control). Pendimethalin (0.9 kg ai ha−1) and S-metolachlor (1.4 kg ha−1) were applied PRE to the entire test. Plots were maintained by hand weeding or applications of graminicides for the entire season to remove all weeds except Florida beggarweed.
In Georgia, final Florida beggarweed populations (plants m−2) were determined in October of each year by counting all plants in each plot. Prior to peanut harvest, five Florida beggarweed plants were randomly hand harvested from each plot by cutting plants at the soil surface, storing them in paper bags, and then drying for 96 hrs using heated (80 C) forced air. Florida beggarweed dry weight was recorded and dry weights were converted to kg ha−1 biomass. In Alabama, only fresh Florida beggarweed weight was determined by hand harvesting the entire plot and converting to kg ha−1. At the Georgia locations, peanut was dug and inverted based on pod mesocarp color (Williams and Drexler, 1981). Pods were combined 10 to 20 d later with conventional harvesting equipment and yield reported on a kg ha−1 basis. Peanut yield was not collected for the Alabama studies.
Data were subjected to mixed models analysis of variance for the eleven (PRE, EPOST, and POST herbicide treatments) by two (LPOST treatments) factorial treatment arrangement. As a result, data for locations were combined across years. Locations were then analyzed separately and means for significant main effects and interactions were separated using Fisher's Protected LSD method test at P ≤ 0.05.
Results and Discussion
For Georgia, the two-way interactions between the eleven PRE, EPOST, and POST treatments and 2 LPOST treatments were not significant for Florida beggarweed population, dry weight, and peanut yield. Therefore, data for the main effects were combined for presentation. The two-way interaction between the eleven PRE, EPOST, and POST herbicide treatments by the two LPOST treatments was significant for Florida beggarweed control in Georgia and Alabama and fresh weight biomass in Alabama.
Florida beggarweed stand and biomass
In Georgia, the population of Florida beggarweed was significantly reduced by the application of any residual herbicide (flumioxazin, diclosulam, or imazapic) alone at any rate either PRE or POST compared to the nontreated control (Table 1). Flumioxazin at 105, diclosulam at 53, or imazapic at 71 g ha−1 reduced Florida beggarweed stand to 0.9, 1.4, and 0.5 plants m−2, respectively, compared to nontreated control with 2.9 plants m−2. Use of the contact treatment of paraquat plus bentazon plus 2,4-DB EPOST alone did not reduce Florida beggarweed populations (2.6 plants m−2) as compared to the nontreated control. The addition of chlorimuron LPOST did not reduce Florida beggarweed population, as noted with no significant interactions (data not shown). These data indicate that the addition of chlorimuron did not reduce Florida beggarweed population but merely provided suppression of existing plants. Grey and Bridges (2005) reported a similar finding in that chlorimuron applied at 49 or 63 days after peanut emergence suppressed Florida beggarweed biomass production but did provide complete control.
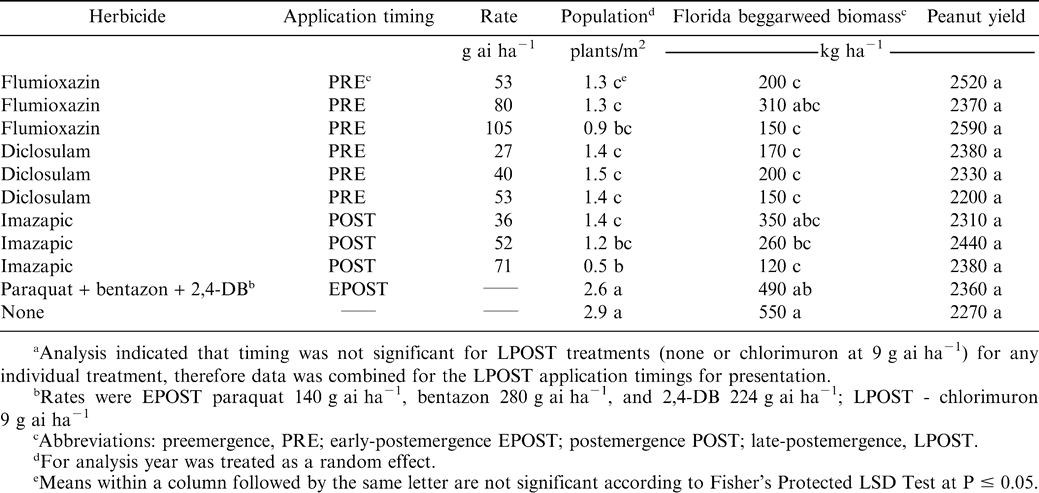
Florida beggarweed population, dry weight biomass, and peanut yield as influenced by herbicide for Ty Ty GAa.
Florida beggarweed dry weight biomasses in Georgia were 150, 150, and 120 kg ha−1 for flumioxazin at 105 PRE, diclosulam at 53 PRE, and imazapic at 71 g ha−1 EPOST, respectively (Table 1). Diclosulam at any rate gave significant reduction in Florida beggarweed dry biomass. In contrast, Florida beggarweed dry biomass for the nontreated control was 550 kg ha−1, which was similar to the paraquat plus bentazon plus 2,4-DB treatment (490 kg ha−1). Flumioxazin at 80 g ha−1 resulted in an increase in Florida beggarweed biomass, which could be attributed to partial control which allowed escaped plants to grow with less competition. In Alabama, when chlorimuron was applied to any treatment, data indicated the chlorimuron did not eliminate Florida beggarweed, but provided suppression of existing plants as indicated by reductions in fresh weight biomass (Table 2). This was more noticeable when initial PRE, EPOST, or POST applications did not give good control, as in imazapic at either 36 or 52 g ha−1 or paraquat plus bentazon plus 2,4-DB.
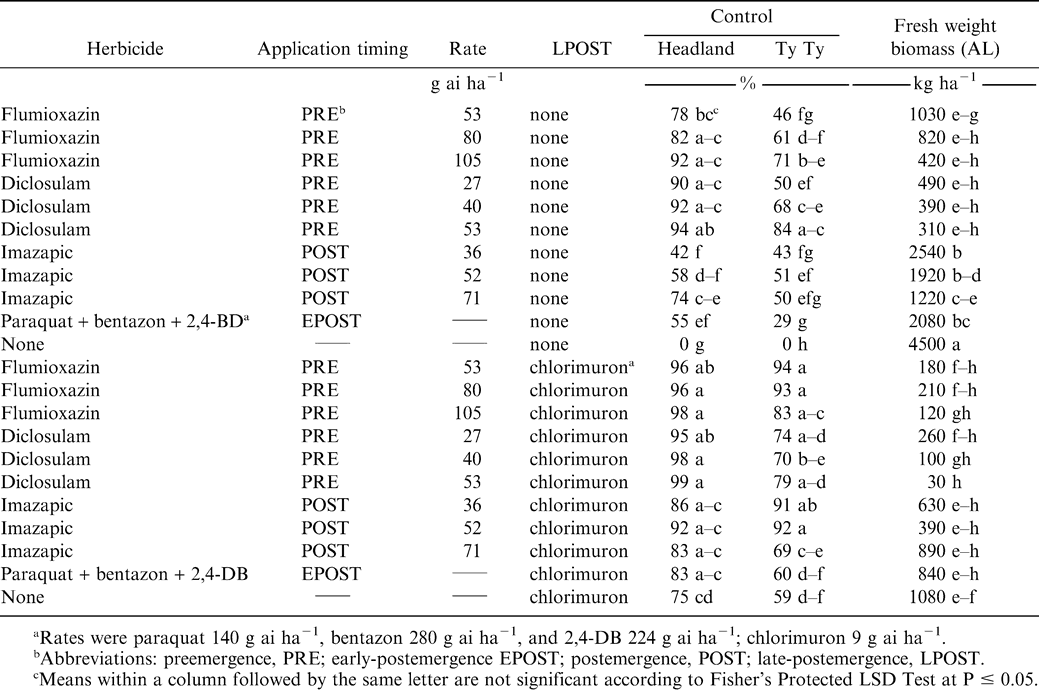
Florida beggarweed control and fresh weight biomass as influenced by herbicide combination for Headland AL and Ty Ty GA.
Florida beggarweed control
While biomass of Florida beggarweed may have been reduced by the use of PRE and POST residual treatments, this did not translate into consistent visual control. All PRE and POST herbicide treatments provided variable Florida beggarweed control (Table 2).
Flumioxazin PRE Florida beggarweed control was 71% or less for any treatment in Georgia. Control was rate dependent for Georgia in 2005 and Alabama. For both locations, 105 g ha−1 was required to obtain maximum levels of control. Previous research has indicated that Florida beggarweed was controlled at least 89% when flumioxazin was PRE applied at 71 to 105 g ha−1 (Grey et al., 2002; 2003). Thus, for maximum Florida beggarweed control, the 105 g ha−1 rate was required. Diclosulam PRE Florida beggarweed control was similar for Georgia in 2005 and 2006 to flumioxazin PRE. However, in Alabama control was 90% or greater for any diclosulam treatment. Across all rates, imazapic POST Florida beggarweed control was 51% or less for Georgia. Florida beggarweed control exhibited a rate response for imazapic POST ranging from 42 to 74% in Alabama. The developing crop canopy can cover Florida beggarweed seedlings and inhibit herbicide spray contact (Cardina and Brecke, 1991). This can limit imazapic POST weed activity resulting in variable Florida beggarweed control which was observed in these and other trials (Grey and Wehtje, 2005). Paraquat plus bentazon plus 2,4-DB Florida beggarweed control was 55% or less across all experiments and has been previously noted (Wehtje et al., 2000a; 2000b).
The addition of chlorimuron LPOST improved Florida beggarweed control for all PRE, EPOST, and POST treatments (Table 2). Generally, the less Florida beggarweed control observed by an early season treatment, the greater the benefit obtained from chlorimuron LPOST. In Alabama, Florida beggarweed control was 95% and greater for PRE treatments of flumioxazin and diclosulam followed by chlorimuron LPOST. Similarly, imazapic followed by chlorimuron Florida beggarweed control was improved to 83% or greater. These Florida beggarweed responses indicate that using either a PPO or ALS herbicide PRE or POST followed by a LPOST ALS application did not reduce Florida beggarweed control.
Florida beggarweed morphology can vary with location and this contributes to its adaptability to changing environments, cultural practices, and control measures (Cardina and Brecke, 1989). Thus, biotype differences between the Alabama and Georgia populations could have resulted in different responses to herbicide treatment. By using multiple applications of the same mode of action, there is an increased potential for selection of ALS herbicide resistant weeds (Retzinger and Mallory-Smith, 1997; Niekamp et al., 1999). Given the adaptability of Florida beggarweed to different environments, there is always potential for development of ALS resistance, especially with the over use of herbicides with that same mode of action. In Georgia the use of ALS herbicides has continued to increase since their introduction. The use of ALS herbicides has increased in peanut from 63% of the hectares treated in 1999 to 93% treated by 2003 (UGA extension survey, 1997–2003).
Peanut yield
Peanut yield in Georgia ranged from 2270 to 2590 kg ha−1 (Table 1). Peanut yield was variable and there was no significant difference between PRE, EPOST, or POST treatments. Overall peanut yield was poor and not indicative of weed control. Buchanan et al. (1976) reported peanut biomass correlated more closely with the weights of the Florida beggarweed rather than with their populations.
Conclusions
Variable Florida beggarweed control (29 to 94%) was noted when flumioxazin PRE, diclosulam PRE, paraquat plus bentazon plus 2,4-DB EPOST, or imazapic POST were applied alone. Florida beggarweed control was 68 to 98% with combinations of flumioxazin PRE at 105 g ha−1, diclosulam PRE at 53 g ha−1, and imazapic POST at 71 g ha−1 followed by chlorimuron LPOST at 9 g ha−1. Florida beggarweed population was not reduced when chlorimuron was LPOST applied following any residual herbicide PRE or POST application. This is of concern since seed would still be produced by these plants. If ALS resistance does develop in Florida beggarweed, it will likely occur as a result of these types of control methods. Weed resistance to ALS herbicides occurs because of the repetitive use of this mode of action in multiple crops, and the ability of different mutations to hinder herbicide binding (McNaughton et al. 2005; Tranel and Wright 2002). For this study, chlorimuron did reduce Florida beggarweed biomass but it did not reduce the populations. Therefore, growers should utilize PRE herbicides that will effectively control Florida beggarweed early in the season and not be reliant on POST applications.
Acknowledgements
The authors wish to thank Charlie Hilton and Darrell Bryner for technical assistance with this research. The authors thank BASF Corp., Dow Agro Sciences Corp. and Valent Corp. for providing materials and assistance with this research.
Literature cited
Anderson R. J. , Norris A. E. , and Hess F. D. 1994 Synthetic organic chemicals that act through to porphyrin pathway. 317 – 342 In Duke S. O. and Rebeiz C. A. Pesticides: Chemistry, toxicology, and pharmaceutical applications. ACS Symposium Series No. 559, American Chemical Society, Washington, DC.
Branch W. D. 1996 Registration of ‘Georgia Green’ peanut. Crop Sci 36 : 806 .
Buchanan G. A. , Murray D. S. , and Hauser E. W. 1982 Weeds and their control in peanuts. 206 – 249 In Pattee H. E. and Young C. T. Peanut Science and Technology, American Peanut Research and Education Society, Inc Yoakum, TX .
Buchanan G. A. , Hauser E. W. , Ethredge W. J. , and Cecil S. R. 1976 Competition of Florida beggarweed and sicklepod with peanuts. II. Effects of cultivation, weeds, and SADH. Weed Sci 24 : 29 – 39 .
Cardina J. and Brecke B. J. 1991 Florida beggarweed (Desmodium tortuosum) growth and development in peanuts (Arachis hypogaea). Weed Technol 5 : 147 – 153 .
Dowler C. C. 1992 Weed survey - southern states. Proc. South. Weed Sci. Soc 45 : 396 (abstr.) .
Dowler C. C. 1998 Weed survey - southern states. Proc. South. Weed Sci. Soc 51 : 304 (abstr.) .
Elmore C. D. 1989 Weed survey - southern states. Proc. South. Weed Sci. Soc 42 : 413 (abstr.) .
Grey T. L. and Bridges D. C. 2005 Control method and time of emergence effects on Florida beggarweed (Desmodium tortuosum) competition in peanut (Arachis hypogaea). Peanut Sci 32 : 73 – 80 .
Grey T. L. , Bridges D. C. , Eastin E. F. , and MacDonald G. E. 2002 Influence of application rate and timing of flumioxazin on weed control in peanut (Arachis hypogaea). Peanut Sci 29 : 24 – 29 .
Grey T. L. , Bridges D. C. , Eastin E. F. , Prostko E. P. , Vencill W. K. , Johnson W. C. , Brecke B. J. , MacDonald G. E. , Dacur J. A. , Everest J. W. , Wehtje G. R. , and Wilcut J. W. 2003 Residual weed control with imazapic, diclosulam, and flumioxazin in southeastern peanut (Arachis hypogaea). Peanut Sci 30 : 23 – 28 .
Grey T. L. and Wehtje G. W. 2005 Residual weed control systems in peanut (Arachis hypogaea). Weed Technol 19 : 560 – 567 .
Hauser E. W. , Buchanan G. A. , and Ethredge W. J. 1975 Competition of Florida beggarweed and sicklepod with peanuts. I. Effects of periods of weed-free maintenance or weed competition. Weed Sci 23 : 368 – 372 .
Hauser E. W. , Buchanan G. A. , and Ethredge W. J. 1982 Effects of Florida beggarweed (Desmodium tortuosum) and sicklepod (Cassia obtusifolia) on peanuts. Weed Sci 30 : 602 – 604 .
Johnson W. C. , Colvin D. L. , Littlefield T. A. , and Mullinix B. G. 1999 Florida beggarweed (Desmodium tortuosum) and sicklepod (Senna obtusifolia) control in peanut using herbicides applied through a wick-bar. Peanut Sci 26 : 18 – 23 .
Johnson W. C. , Holbrook C. C. , Mullinix B. G. , and Cardina J. 1992a Response of eight genetically diverse peanut genotypes to chlorimuron. Peanut Sci 19 : 111 – 115 .
Johnson W. C. , Mullinix B. G. , and Brown S. M. 1992b Phytotoxicity of chlorimuron and tank mixtures on peanut (Arachis hypogaea). Weed Technol 6 : 404 – 408 .
McNaughton K. E. , Letarte J. , Lee E. A. , and Tardiff F. J. 2005 Mutations in ALS confer herbicide resistance in redroot pigweed (Amaranthus retroflexus) and powell amaranth (Amaranthus powellii). Weed Sci 53 : 17 – 22 .
National Agricultural Statistics Service (NASS) 2004 National Agricultural Statistics Service U.S. Dept. Agri., Published Estimates Database. NASS-USDA, Washington, DC. Online at http://www.pestmanagement.info/nass/. Accessed on August 27, 2007.
Niekamp J. W. , Johnson W. G. , and Smeda R. J. 1999 Broadleaf weed control with sulfentrazone and flumioxazin in no-tillage soybean (Glycine max). Weed Technol 13 : 233 – 238 .
Prostko E. P. 2001 Weed survey - southern states. Proc. South. Weed Sci. Soc 54 : 249 (abstr.).
Retzinger E. J. and Mallory-Smith C. M. 1997 Classification of herbicides by site of action for weed resistance management strategies. Weed Technol 11 : 384 – 391 .
Tranel P. J. and Wright T. R. 2002 Resistance of weeds to ALS-inhibiting herbicides: what have we learned? Weed Sci 50 : 700 – 712 .
Vencill W. K. 2002 Herbicide Handbook. 8th ed Weed Sci Soc of America Lawrence, KS .
Webster T. M. 2005 Weed survey - southern states. Proc. South. Weed Sci. Soc 58 : 291 – 306 (abstr.) .
Webster T. M. and MacDonald G. E. 2001 A survey of weeds in various crops in Georgia. Weed Technol 15 : 771 – 790 .
Webster T. M. , Wilcut J. W. , and Coble H. D. 1997 Influence of AC 263,222 rate and application method on weed management in peanut (Arachis hypogaea). Weed Technol 11 : 520 – 526 .
Wehtje G. W. and Grey T. L. 2005 Response of selected peanut (Arachis hypogaea) cultivars to early postemergence chlorimuron applications. Peanut Sci 32 : 119 – 123 .
Wehtje G. R. , McGuire J. A. , Walker R. H. , and Paterson M. G. 1986 Texas panicum control in peanuts with paraquat. Weed Sci 34 : 308 – 311 .
Wehtje G. R. , Padgett D. , and Martin N. R. 2000a Performance and economic benefit of herbicides used for broadleaf weed control in peanut. Peanut Sci 27 : 11 – 16 .
Wehtje G. R. , Padgett D. , and Martin N. R. 2000b Imazapic-based herbicide systems for peanut and factors affecting activity on Florida beggarweed. Peanut Sci 27 : 17 – 22 .
Wehtje G. R. , Wells L. W. , Choate J. H. , Martin N. R. , and Curtis J. M. 1999 Mowing as a weed control supplement to herbicides and cultivation in peanut (Arachis hypogaea). Weed Technol 13 : 139 – 143 .
Wilcut J. W. , York A. C. , Grichar W. J. , and Wehtje G. R. 1995 The biology and management of weeds in peanut (Arachis hypogaea). 207 – 244 In Pattee H. E. and Stalker H. T. Advances in Peanut Science American Peanut Research and Education Society, Inc Stillwater, OK.
Williams E. J. and Drexler J. S. 1981 A non-destructive method for determining peanut pod maturity. Peanut Sci 8 : 134 – 141 .
Wise A. M. , Grey T. L. , Prostko E. P. , Vencill W. K. , and Webster T. M. 2009 Establishing the geographic distribution level of acetolactate synthase resistance of Palmer amaranth (Amaranthus palmeri) accessions in Georgia. Weed Technol 23 . (In press).
Yoshida M. , Sakaki M. R. , Sato R. , Haga T. , and Kamoshita K. 1991 S-53482 a new N-phenyl-pthalimide herbicide, in Proc. Brighton Crop Prot. Conf.: Weeds, 69 – 75 (abstr.).
Notes
Author Affiliations
1 Assoc. Prof. and Assoc. Prof, Dept. of Crop and Soil Sci., respectively, The University of Georgia, Tifton Campus, P.O. Box 748, 115 Coastal Way, Tifton, GA 31793.
2 Prof., Agronomy Dept., Auburn University, Auburn AL 36849, respectively.
*Corresponding author (email: tgrey@uga.edu).