

Introduction
Flavor is among the most important quality attributes for commercial acceptance of roasted peanuts (Arachis hypogaea L.) as snacks or peanut butter. Manufacturers of peanut products desire to deliver consistently flavorful products. Peanut breeding programs are sensitive to the need to at least maintain, but ideally improve, peanut flavor of new cultivars. However, peanut flavor is influenced by several groups of factors: environmental factors (associated with years, production regions, locations within regions, and interactions of region and location with year), genetic factors (cultivars), and interaction between environmental and genetic factors (genotype-by-environment or “G×E” interaction). In order to ascertain reasonable goals with respect to consistency of flavor of peanut products, one must know the relative contributions of genotype, environment, and G×E interaction to sensory quality.
In 2002, the USDA Agricultural Research Service (USDA-ARS) implemented a sensory quality testing program for peanuts grown as part of the Uniform Peanut Performance Test (UPPT) (Branch et al., 2006). The UPPT is a collaborative program in which advanced breeding lines developed by the several public-sector peanut breeding programs are evaluated for agronomic performance and grade across the three US peanut-producing regions. Beginning in 2002, USDA-ARS personnel applied standard sensory evaluation procedures to samples of UPPT entries from each test site. The UPPT flavor data set provides a unique opportunity to examine the contributions of genotype, environment, and G×E interaction to variation in sensory quality. Previous estimates of variance components, broad-sense heritabilities, and standard errors of the differences between means were based on analysis of data collected on samples donated by breeding and variety testing programs in different states (Pattee and Giesbrecht, 1990; Pattee et al., 1993, 1994, 1997, 1998). Although samples of Florunner, NC 7, or a spanish or Valencia market-type check were submitted along with samples of experimental breeding lines, the data collected on donated samples suffers from a lack of orthogonality. In contrast, each year's UPPT includes 13 to 16 breeding lines and cultivars evaluated at each of 9 locations across 7 states. Within years, there is perfect orthogonality. Experimental UPPT entries generally do not stay in the test for more than two years, but the check cultivars Florunner and NC 7 are included in all years, providing greater across-year orthogonality than is found in the donated data set. The objective of this study was to use published UPPT results to estimate variance components associated with environmental, genotypic, and G×E effects on sensory quality of peanuts.
Materials and Methods
Methodology of the USDA-ARS Market Quality and Handling Research Unit (MQHRU) at Raleigh, NC has been published in annual reports available online (Uniform Peanut Performance Tests, 2003. Chemical, Sensory and Shelf-life Properties. Data Presented by Location. July, 2003. USDA-ARS-SAA Market Quality and Handling Research Unit, Raleigh, NC, pp. 4–7, http://152.1.118.33/, validated 21 Feb 2007). The data from the UPPT include paste color and sensory attribute intensities measured on bulk samples from the replicate plots grown at each of nine test locations (Suffolk, VA and Lewiston, NC in the Virginia-Carolina production area; Tifton, GA, Marianna, FL, and Headland, AL in the Southeastern production area; and Denver City or Brownfield, TX, Stephenville, TX, Pearsall, TX, and Fort Cobb, OK in the Southwestern production area). Processing of the shelled peanuts resulted in samples separated into US medium and jumbo grade size fractions for runner types. Whenever possible, both fractions were subjected to sensory analysis. For virginia market-type entries, only the extra large kernel fraction was analyzed.
Data from the 2002 through 2005 crop years were used in the analysis. The mixed models procedure (PROC MIXED) of the SAS statistical software package (SAS Inst., Cary, NC) was used to apply restricted maximum likelihood (REML) estimation of variance components associated with year, production region, year-by-region interaction, location within regions, year-by-location interaction in region, genotype, year-by-genotype interaction, region-by-genotype interaction, year-by-region-by-genotype interaction, location-by-genotype interaction in region, year-by-location-by-genotype interaction in region, kernel grade within genotype, year-by-grade interaction in genotype, region-by-grade interaction in genotype, year-by-region-by-grade interaction in genotype, location-by-grade interaction in region and genotype. All of these effects were considered random in order to estimate a variance component. All other effects were pooled as a residual effect.
Repeatability coefficients were estimated as indicators of the magnitude of the genetic component of variance relative to the variance among genotype means estimated from one or two years in the UPPT trials. The estimates were obtained as
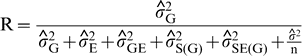
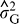 is the variance attributable to genotypes,
is the variance attributable to genotypes,
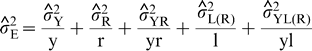 is the estimate of variance attributable to environments,
is the estimate of variance attributable to environments,
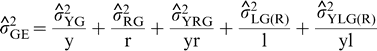 is the estimate of variance attributable to G×E interaction,
is the estimate of variance attributable to G×E interaction,
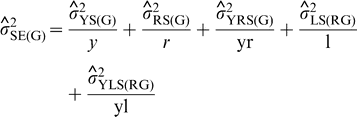
Results and Discussion
Roasted peanut is the most important among the sensory attributes. It exhibited very little genetic variation, approximately 1% of the total (Table 1). This is substantially less than the estimated proportion (6 to 31%) published previously (Pattee et al., 1990, 1993, 1995, 1998). The highest estimate was obtained from data collected only on virginia-type lines grown at two fairly close locations (Suffolk, VA and Pendleton, NC), perhaps restricting the amount of environmental and G×E variation and thereby increasing the relative proportion due to genetic variation. The lowest and most recent estimate (Pattee et al., 1998) was calculated from data on samples including all four market types of peanuts and representing environments from three production regions and several years. The wider genetic representation should have maximized the genetic variance, but the greater environmental representation should have increased the environmental and G×E components. In the current study, UPPT entries were either runner or virginia market-types, perhaps constraining genetic variance, while the environmental representation was wide. Environmental effects accounted for 32% of the total variation for roasted peanut, and G×E effects 45%, almost all of it year-by-genotype (Y×G) interaction. The form of G×E effect previously found to have the largest impact on the roasted peanut attribute was between genotypes and specific locations within years and regions (Pattee et al., 1998). In that study, Y×G interaction was a minor contributor to the overall G×E variation. The relatively small influence of Y×G interaction in previous studies may reflect the limited number of genotypes replicated across years in those studies. Although the number of UPPT entries retained from year to year is limited, there are almost always a few experimental lines retained for a second or greater year, and the checks Florunner and NC 7 are included every year at all locations. Therefore, the UPPT data should provide a better estimate of the Y×G component of variance than was previously available.
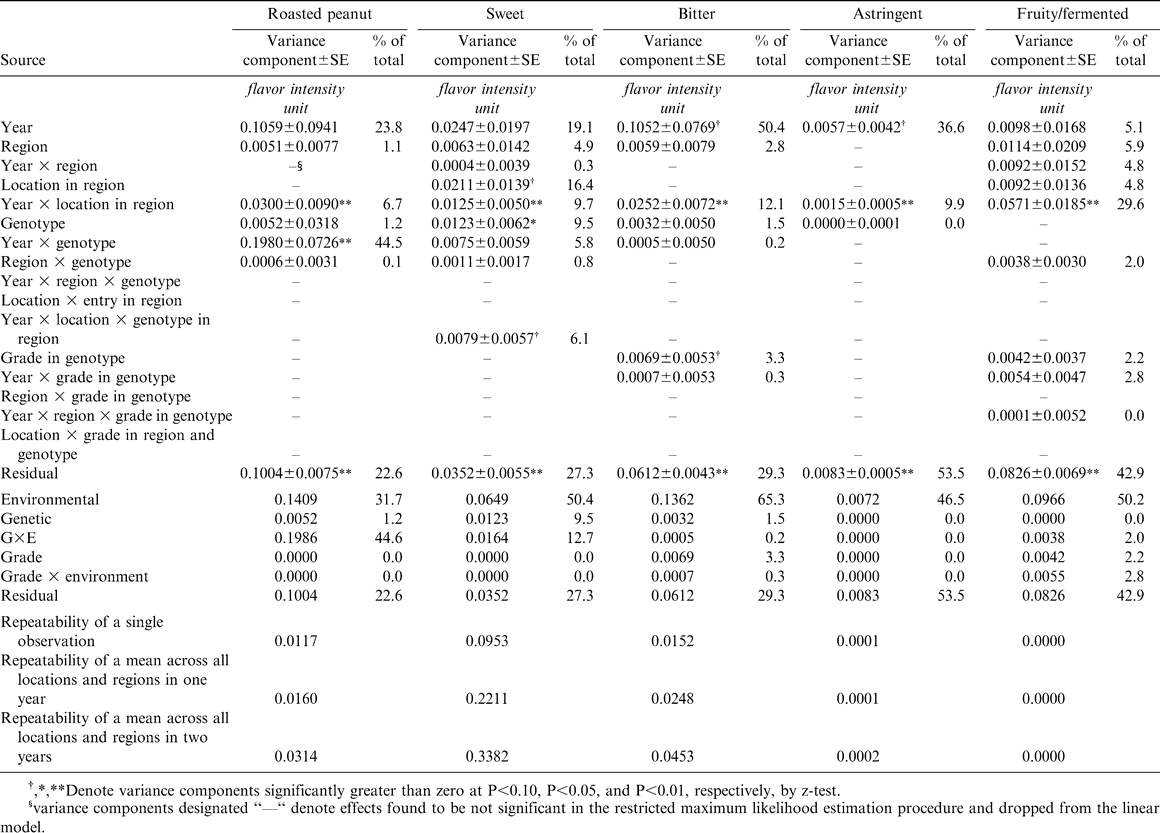
Variance component estimates for the roasted peanut, sweet, bitter, astringent, and fruity/fermented sensory attribute intensities of processed peanut samples from the 2002–2005 Uniform Peanut Performance Test.
For the sweet attribute, genotypes accounted for the largest proportion of total variation of any sensory attribute, 9.5% (Table 1). Likewise, the relative magnitude of environmental factors (50.4% of total variation) was larger than that for roasted peanut although the absolute magnitude of the environmental variance component for sweet was approximately half that for roasted peanut. However, G×E variance was much smaller for sweet than for roasted peanut both in absolute value and relative to total variance. The sweet attribute had the greatest repeatability coefficients of any of the sensory attributes whether considering a single observation (R = 0.10) or means across one (R = 0.22) or two years (R = 0.34) of UPPT testing. Repeatability of two-year means for roasted peanut were estimated at only R = 0.03. In a practical sense, this implies that breeders should be able to identify breeding lines with superior sweet scores with a high level of confidence based on means across one or two years of multiple-year testing, but that lines with superior roasted peanut intensity will be more difficult to identify accurately.
The bitter attribute exhibited the most environmental variation as a proportion of total of any sensory attribute even though the actual estimate of the environmental component of variance was very close to that for roasted peanut (0.1362 versus 0.1409 flavor intensity units or “fiu”) (Table 1). As was the case for roasted peanut, most of the environmental variation for bitter was associated with years rather than spatial factors such as production region or location within region. There was very little G×E variation for bitter. Repeatability of estimates of bitter was very similar to that for roasted peanut.
Genetic effects accounted for only a small proportion of each of the other sensory attributes (Tables 1 and 2), ranging from 0% of total variation for the fruity/fermented, astringent, and raw/beany attributes to 4.6% of the sweet aromatic attribute. Environmental effects were a much larger source of variation for sensory quality (24.5 to 50.2% of total variation). Like the roasted peanut attribute, the sweet aromatic attribute exhibited a large effect of Y×G interaction.
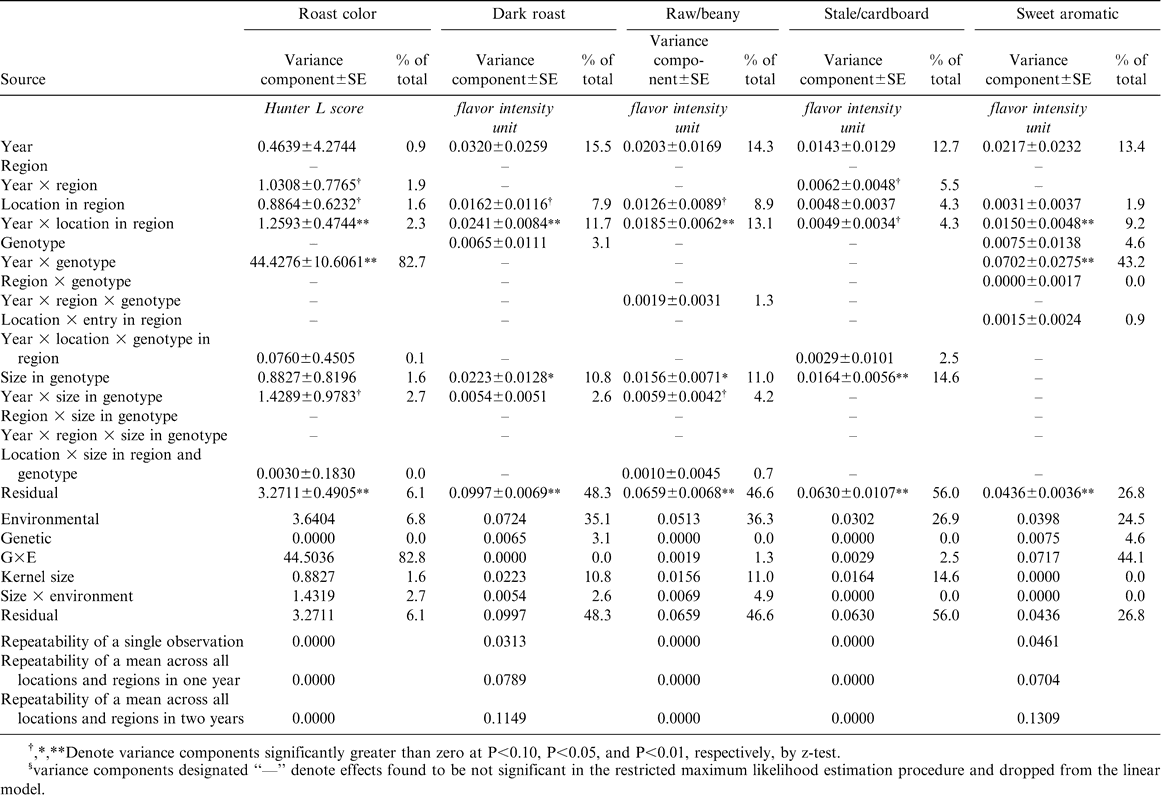
Variance component estimates for roast color and the dark roast, raw/beany, stale/cardboard, and sweet aromatic attribute intensities of processed peanut samples from the 2002–2005 Uniform Peanut Performance Test.
Of particular note is the magnitude of environmental effects for the fruity/fermented attribute, 50% of the total, with the balance due to the effects of sample-to-sample (experimental) error. Among the environmental effects on the fruity/fermented attribute, specific combinations of years and locations within regions were the largest (30% of total variation). This sort of effect specific to a particular year at a particular location may reflect the average maturity of the peanuts at harvest, or it might reflect a problem with post-harvest handling of the pod samples from a specific UPPT trial.
Roast color exhibited no variation associated with genotypes. Given that the roasting protocol called for each sample to be roasted to a degree that would result in a common paste darkness (Hunter L score of approximately 49), one might expect there to be no variation in roast color. However, there was a small environmental influence on roast color and a substantial G×E variance (82.8% of total variation), again due primarily to Y×G effects. Roast color is often associated with the two sensory attributes dark roast and raw/beany. Peanuts roasted too light tend to have higher raw/beany intensity while peanuts roasted too dark have greater dark roast intensity. These two sensory attributes exhibit strong negative correlation (data not shown), and the estimates of variance components for the two attributes in the current study mirror each other as one might expect, but they do not correlate with the variance components for roast color. Raw/beany and dark roast exhibited very little variation that could be attributed to design effects in the linear statistical model describing the UPPT trials. Almost half of the total variation was associated with random effects usually called experimental error or “noise.”
The preponderance of environmental influence on sensory quality of peanuts raises the question of which specific environmental factors produce the variation. The relatively large effect of years on flavor suggests factors that affect the entire peanut-growing area of the U.S.A. Variation in temperature and rainfall tends to be more localized than the entire peanut-growing area, so it does not seem likely that these are the factors of interest. Other potential factors include changes in the disease and insect complexes that affect peanut growth, in the agrichemicals applied to manage diseases and pests, or other cultural practices such as planting dates, harvest dates, rotations, and drying protocols.
Literature Cited
Branch W. D. , Coker D. L. , Isleib T. G. , Chapin J. W. , Bostick J. P. , Gorbet D. W. , Tillman B. L. , Simpson C. E. , Burow M. D. , Baring M. L. , and Greenhagen B. 2006 Uniform Peanut Performance Tests, 2005. Coastal Plain Exp. Sta. Res. Prog. Rep. No. 4-06. 25 pp.
Pattee H. E. and Giesbrecht F. G. 1990 Roasted peanut flavor variation across germplasm sources. Peanut Sci 17 : 109 – 112 .
Pattee H. E. , Giesbrecht F. G. , and Mozingo R. W. 1993 A note on broad-sense heritability of selected sensory descriptors in virginia-type Arachis hypogaea L. Peanut Sci 20 : 20 – 24 .
Pattee H. E. , Isleib T. G. , and Giesbrecht F. G. 1994 Genotype-by-environment interaction in roasted peanut attribute. Peanut Sci 21 : 94 – 99 .
Pattee H. E. , Giesbrecht F. G. , and Isleib T. G. 1995 Roasted peanut flavor variation among U.S. peanut genotypes. Peanut Sci 22 : 158 – 162 .
Pattee H. E. , Isleib T. G. , and Giesbrecht F. G. 1997 Genotype-by-environment interaction in the sweet and bitter attributes of peanut. Peanut Sci 24 : 117 – 123 .
Pattee H. E. , Isleib T. G. , and Giesbrecht F. G. 1998 Variation in intensity of sweet and bitter sensory attributes across peanut genotypes. Peanut Sci 25 : 63 – 69 .
Notes
- First and second authors: Res. Agron. , USDA-ARS and Agric. Res. Statistician; Coastal Plain Exp. Sta., Tifton, GA 31793-0748. Third author: President and CEO, Hebert Green AgroEcology; Asheville, NC 28801. [^]
- T.G. Isleib, Dep. of Crop Sci., Box 7629, N.C. State Univ., Raleigh, NC 27695-7629. [^]
- B.L. Tillman, Univ. of Fla. N. Fla. Res. & Ext. Ctr., 3925 Hwy. 71, Marianna, FL 32446-8091. [^]
- H.E. Pattee, Dep. of Biol. & Agric. Engineering, Box 7625, N.C. State Univ., Raleigh, NC 27695-7625. [^]
- T.H. Sanders, K.W. Hendricks, and L.O. Dean, USDA-ARS Market Quality and Handling Res. Unit, Raleigh, NC 27695-7624. [^] * Corresponding author. E-mail: tom_isleib@ncsu.edu.
Author Affiliations