

Introduction
Improvement of peanut sensory quality is a goal of the peanut breeding program at N.C. State Univ. It has been documented that the intensity of the roasted peanut sensory attribute in peanuts of the virginia market-type is, on average, weaker than its intensity in runner-type peanuts, although the distributions of roasted peanut intensity across cultivars and breeding lines of the two market types were found to overlap (Isleib et al., 2001; Pattee et al., 1994, 1995, 1997, 1998). Similarly, runner-type peanuts have superior average scores for the sweet and bitter attributes. Best linear unbiased prediction (BLUP) has been used to identify peanut germplasm with putatively beneficial breeding value for sensory attributes roasted peanut, sweet, and bitter (Pattee et al., 2001, 2002a, 2003), but the practical utility of those predictors of breeding value has not been validated.
The objective of this study was to empirically estimate the combining abilities of parents for sensory quality and to compare those combining abilities with BLUPs and with mean performance of the parents as lines per se.
Materials and Methods
Mating Design
Eleven cultivars and breeding lines were chosen for use as parents of a half-diallel cross. Nine of the parents were selected on the basis of BLUPs of breeding value for the roasted peanut, sweet, and bitter sensory attributes (Pattee et al., 2001, 2002a); they included two runner-type lines [F439-2-3-2-1, a sibling of Florunner (Norden et al., 1969), and Andru 93 (Gorbet and Knauft, 1995)], four virginia-types [Florida breeding line UF714021, a component of the Altika cultivar (Norden and Gorbet, 1974), NCSU breeding lines N97054J selected from cross X90053 (Pattee et al., 2002b) and NC Ac 18457, and landrace selection White's Runner], two spanish-type cultivars [Pearl and Pronto (Banks and Kirby, 1983)], and one valencia-type cultivar [New Mexico Valencia C (Hsi, 1980)]. In addition to the parents predicted to have positive effects on peanut flavor, two additional parents were included because of their widespread use among peanut growers: runner-type cultivar Georgia Green (Branch, 1996) and virginia-type cultivar Perry ( Isleib et al., 2003). Georgia Green was later found to have positive BLUPs of breeding value for flavor (Pattee et al., 2003) while Perry was a cultivar with negative BLUPs (Pattee et al., 2001, 2002a). The 11 parents were crossed in a half-diallel mating in the summer of 2001 in the greenhouse at the NCSU campus in Raleigh, NC. Each parent was used five times as a male and five as a female in the mating design.
Field Procedures
F1 hybrids were grown in a winter nursery at Juana Diaz, PR, in the winter of 2001–2002. F2 populations were grown at the Peanut Belt Research Station (PBRS) in Lewiston, NC, in 2002, and single plant selections were made on the basis of pod characteristics and maturity as evidenced by bright hull color after the inverted peanuts had been allowed to dry in the windrow for several days. Selections were made in the F3 at PBRS in 2003. F3∶4 families from the same cross were bulked in equal numbers to provide seed for testing in a replicated trial at PBRS in 2004. Thirty-three crosses survived the inbreeding and selection program; these were grown with check cultivars Florunner, NC 7 (Wynne et al., 1979) and Gregory (Isleib et al., 1999) in a randomized complete block design with two replications. Each plot comprised two rows 7.3 m in length spaced 91 cm apart. Seed spacing at planting was 25 cm. Plots were planted on 11 May 2004, dug 1 October 2004, and harvested by combine on 7 October 2004.
Sample Handling, Roasting and Preparation
Pods were sized on a rolling grader and shelled with a standard reciprocating grading sheller (Georgia Federal-State Inspection Service, Albany, GA). Seeds were screened over a 6.0 × 25.4 mm (15/64 × 1 in) grading screen. The shelled peanuts were stored at 5 C and 60% RH. Samples were roasted in May using a Blue M “Power-O-Matic 60” laboratory oven, ground into a paste, and stored in glass jars at −20 C until evaluated. The roasting, grinding, and color measurement protocols were as described by Pattee and Giesbrecht (1990). Roast color of the peanut paste was evaluated as CIELAB L*.
Sensory Evaluation
An eight-member, trained, roasted peanut profile panel at the Food Science Dept. at NCSU, Raleigh, NC, conducted a descriptive sensory analysis of all peanut-paste samples using a 14-point intensity scale. Panel orientation and reference control were as described by Pattee and Giesbrecht (1990) and Pattee et al. (1993). Sensory evaluation commenced on 7 October 2005 and continued until all samples were evaluated. The averages of individual panelists' scores on sensory attributes were used in all analyses in this study.
Statistical Analysis
Data from the field experiment were analyzed using the general linear models procedure (PROC GLM) of the SAS statistical software package (SAS Institute, Cary, NC). A design matrix was developed to partition the total variation among the 36 entries in the trial into parts due to (1) the difference between the three checks and the hybrid populations, (2) variation among the three checks with one contrast between Florunner and the two virginia checks and a second between NC 7 and Gregory, and (3) variation among the hybrid populations based on Griffing's (1956) Model 1 Method 1. All 11 parents were represented in the hybrid populations, and general combining ability (GCA) was estimable for all 11. Because only 33 of the 55 crosses survived the inbreeding and selection phase of population development, not all specific combining ability (SCA) effects were estimable. Those effects that were not estimable were assumed to be zero, and the remaining SCA effects were left in the model. When variation due to SCA was not significant, the model was run without SCA effects to allow estimation of GCA effects without the distortion that arises from confounding of SCA with GCA effects in the unbalanced set of crosses. GCA estimates were separated by t-test. The need for use of the covariates fruity and roast color (linear and quadratic) to control error variation was tested for each response variable. Covariates were retained in the model if their effects were significant. If not, they were removed from the model. The error mean square from the full model was used in all tests of significance and in calculating standard errors of effects.
Mean values for intensities of the roasted peanut, sweet, and bitter attributes for the 11 parents were calculated from the same set of data used to calculate BLUPs of breeding value (Pattee et al., 2003). Means were adjusted to common year and location effects and also for the effects of the covariates roast color (linear and quadratic) as an indicator of degree of roast and intensity of the fruity attribute, an indicator of poor environmental conditions pre- and post-harvest. Simple correlations were calculated between GCAs and BLUPs and between GCAs and means.
Results and Discussion
The covariates fruity and roast color were not retained for error control in either roasted peanut or bitter attribute, but they were retained for sweet (Table 1). There was significant variation among test entries for the roasted peanut and sweet attributes, but not for bitter. The difference between the mean of the hybrid populations and that of the checks (Florunner, NC 7 and Gregory) was not significant for any sensory attribute, but there was variation among checks for roasted peanut and sweet. For roasted peanut, the variation arose from the difference between Florunner and the mean of the two virginia-type checks (5.11 vs. 3.88 fiu, P<0.01) and not from the difference between NC 7 and Gregory (3.69 vs. 4.06 fiu, ns), while for sweet the Florunner differed from the virginia-type checks (4.19 vs. 3.27 fiu, P<0.05) and NC 7 differed from Gregory (2.93 vs. 3.61 fiu, P<0.05). Variation among crosses was significant for roasted peanut and sweet with GCA significant for both and SCA for sweet only. SCA usually is considered to be associated with non-additive types of genetic effects, i.e., dominance or epistasis. Because the influence of dominance effects on hybrid means is reduced by half with each generation of self fertilization, it would require very large dominance effects to persist into the F4 generation, suggesting that the persistent non-additive variation detected for the sweet sensory attribute arose due to epistasis (Isleib et al., 1978). GCA effects usually are considered to be indicative of additive genetic effects.
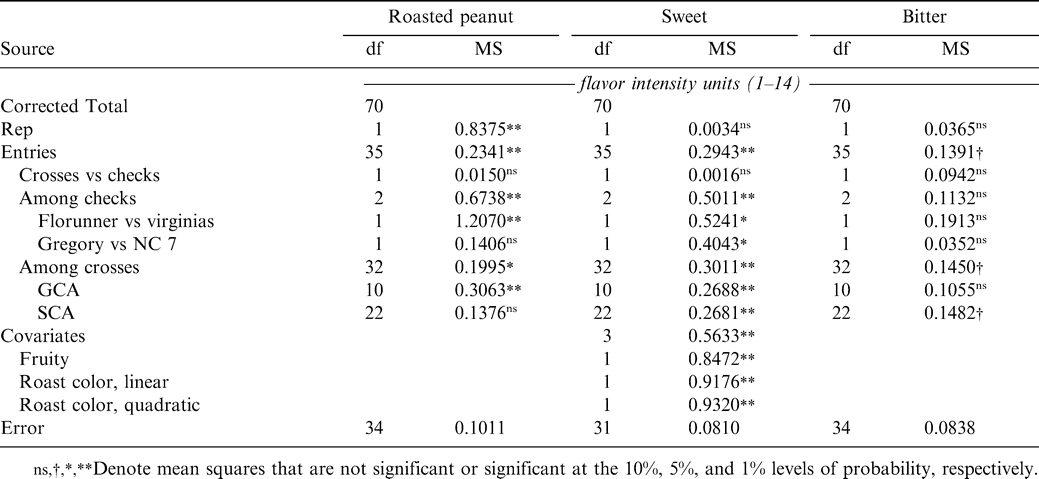
Mean squares from analysis of variance of sensory attribute intensity scores from a diallel mating among 11 parents tested with three checks.
GCA estimates (Table 2) identify UF714021 as the best parent for improvement of roasted peanut and bitter attribute intensities and Pearl for improvement of sweet, although there are several parents not significantly different from these for each of the attributes. Correlations of the empirically determined GCAs with the previously predicted breeding values (Pattee et al., 2003) were positive but moderate in magnitude (r = 0.355, ns for roasted peanut; r = 0.602, P<0.05 for sweet, and r = 0.458, ns for bitter). The corresponding correlations of GCAs with adjusted means calculated from the database used in calculating the BLUPs of breeding value were 0.423 (ns) for roasted peanut, 0.522 (ns) for sweet, and 0.334 (ns) for bitter.
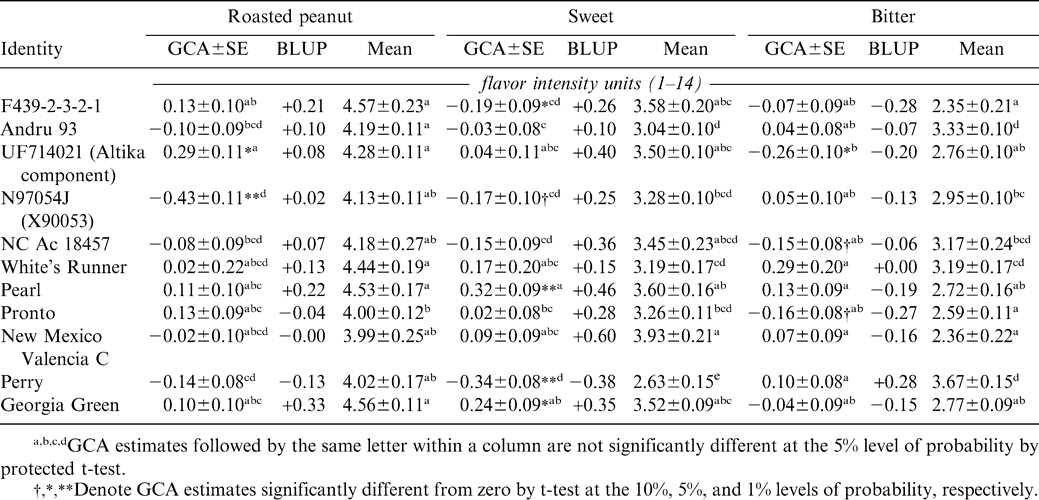
General combining abilities (GCAs) estimated from field study conducted in 2004, best linear unbiased predictors (BLUPs), and means estimated from multiple-year database.
It is not clear whether BLUPs are as useful as empirically derived estimates of combining ability for predicting usefulness of a particular line as a parent in a crossing program designed to improve flavor, nor is it clear that BLUPs were superior to adjusted means as indicators of parental value. However, empirical estimation of GCA for large numbers of potential parents would be impractical due to the cost of the necessarily very large hybridization program and of the sensory analysis of large numbers of samples. BLUPs and adjusted means can be calculated from data collected in ongoing variety tests and the data on different lines need not be collected in the same years and locations. In other words, estimation of GCA requires formal mating designs and large replicated trials of hybrid populations while calculation of BLUPs and means requires only data that will be collected on released and candidate lines as a matter of course. While it may appear that there is no advantage to the use of BLUPs versus means to provide a first approximation of breeding value, it must be recognized that the main additional requirement for calculation of BLUPS is knowledge of the coancestries among the lines whose breeding values are being predicted. This knowledge is essential at a later stage in a flavor improvement program, namely in choosing exactly which parents to cross. If the coancestry of two parents is high, i.e., if they are closely related, then one would not expect much genetic variation within the hybrid population produced by their cross, nor would one expect to realize much genetic gain from selection within the population.
Literature Cited
Banks D. J. and Kirby J. S. 1983 Registration of Pronto peanut (Reg. No. 28). Crop Sci 23 : 184 .
Branch W. D. 1996 Registration of ‘Georgia Green’ peanut. Crop Sci 36 : 806 .
Gorbet D. W. and Knauft D. A. 1995 Registration of ‘ANDRU 93’ peanut. Crop Sci 35 : 1507 .
Griffing B. 1956 Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Boil Sci 9 : 463 – 493 .
Hsi D. C. H. 1980 Registration of ‘New Mexico Valencia C’ peanut. Crop Sci 20 : 113 .
Isleib T. G. , Pattee H. E. , and Giesbrecht F. G. 2003 Narrow-sense heritability of selected sensory descriptors in virginia-type peanuts (Arachis hypogaea L.). Peanut Sci 30 : 64 – 66 .
Isleib T. G. , Pattee H. E. , Gorbet D. W. , and Giesbrecht F. G. 2001 Genotypic variation in roasted peanut flavor quality across 60 years of breeding. Peanut Sci 29 : 92 – 98 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Mozingo R. W. , Bailey J. E. , and Pattee H. E. 2003 Registration of ‘Perry’ peanut. Crop Sci 43 : 739 – 740 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Mozingo R. W. , and Pattee H. E. 1999 Registration of ‘Gregory’ peanut. Crop Sci 39 : 1526 .
Isleib T. G. , Wynne J. C. , and Rawlings J. O. 1978 Estimates of epistasis for diverse peanut cultivars. Peanut Sci 5 : 106 – 108 .
Norden A. J. and Gorbet D. W. 1974 Registration of Altika peanuts (Reg. No. 18). Crop Sci 14 : 339 .
Norden A. J. , Lipscomb R. W. , and Carver W. A. 1969 Registration of Florunner peanuts (Reg. No. 2). Crop Sci 9 : 850 .
Pattee H. E. and Giesbrecht F. G. 1990 Roasted peanut flavor variation across germplasm sources. Peanut Sci 17 : 109 – 112 .
Pattee H. E. , Giesbrecht F. G. , and Mozingo R. W. 1993 A note on broad-sense heritability of selected sensory descriptors in virginia-type Arachis hypogaea L. Peanut Sci 20 : 24 – 26 .
Pattee H. E. , Giesbrecht F. G. , and Isleib T. G. 1995 Roasted peanut flavor intensity variations among U.S. peanut genotypes. Peanut Sci 22 : 158 – 162 .
Pattee H. E. , Giesbrecht F. G. , and Isleib T. G. 1997 Genotype-by-environment interaction in sweet and bitter sensory attributes of peanut. Peanut Sci 24 : 117 – 123 .
Pattee H. E. , Isleib T. G. , and Giesbrecht F. G. 1994 Genotype-by-environment interaction in roasted peanut attribute. Peanut Sci 21 : 94 – 99 .
Pattee H. E. , Isleib T. G. , and Giesbrecht F. G. 1998 Variation in intensity of sweet and bitter sensory attributes across peanut genotypes. Peanut Sci 25 : 63 – 69 .
Pattee H. E. , Isleib T. G. , Giesbrecht F. G. , and Cui Z. 2002a Prediction of parental genetic compatibility to enhance flavor attributes of peanuts. Ch. 17. 217 – 230 In Rajasekaran K. K. , Jacks T. J. , and Finley J. W. Crop Biotechnology, ACS Symposium Series Washington, DC American Chemical Society 829 .
Pattee H. E. , Isleib T. G. , Gorbet D. W. , and Giesbrecht F. G. 2002b Selection of alternative genetic sources of large seed size in virginia-type peanut - Evaluation of sensory, composition and agronomic characteristics. J. Agric. Food Chem 50 : 4885 – 4889 .
Pattee H. E. , Isleib T. G. , Gorbet D. W. , Giesbrecht F. G. , and Cui Z. 2001 Parent selection for roasted peanut flavor quality. Peanut Sci 28 : 51 – 58 .
Pattee H. E. , Isleib T. G. , Gorbet D. W. , Moore K. M. , Lopez Y. , Baring M. R. , and Simpson C. E. 2003 Sensory quality traits of the runner-type peanut cultivar Georgia Green and its value as a parent compared with Florunner. Peanut Sci 30 : 85 – 88 .
Wynne J. C. , Mozingo R. W. , and Emery D. A. 1979 Registration of ‘NC 7’ peanut. Crop Sci 19 : 563 .