

Introduction
Peanut (Arachis hypogaea L.) yield is a product of a complex cascade of reproductive processes both above and below ground starting at approximately three weeks after seed germination. Pod number and eventual crop yield is determined by the number of floral buds that form flowers, the number of flowers that are fertilized and produce pegs, and the number of pegs that successfully penetrate the soil surface and produce pods (Vara Prasad et al., 1999b). Aside from the genetic programming of these processes, climatic conditions and field management techniques greatly influence the success of each step in the process. Abiotic or biotic stress at any point in this process can affect flowering, pegging, and pod yield (Vara Prasad et al., 1999b).
The positive impact of conservation tillage, strip-tillage in particular, on infiltration, runoff and soil quality has resulted in an increase in grower adoption over the last decade (CTIC, 2004). Conservation tillage systems retain at least 30% plant residue on the soil surface between successive crops (Phillips et al., 1980). Benefits of conservation tillage in the production of peanut and other crops are numerous and include: decreased soil erosion and compaction (Merrill et al., 1999; Raper et al., 2000; Yu et al., 2000); decreased soil evaporative losses (Blevins et al., 1971; Reeves 1997); increased soil organic matter (Reeves, 1997); increased soil water infiltration and plant available water (Blevins et al., 1971; Gallaher, 1977); and decreased disease incidence, specifically Tomato spotted wilt virus in peanut (Marois and Wright, 2003; Cantonwine et al., 2006). Despite these advantages, many peanut growers have expressed concern over possible yield decreases associated with conservation tillage, and cite the purported interference of pegging by either the preceding crop stubble or cover crop residues and the possible increased digging losses in conservation systems as the primary reasons behind their apprehension (Grichar and Boswell, 1987; Jordan et al., 2003; Monfort et al., 2004). In part because of these concerns, less than 30% of peanut production utilizes conservation tillage, despite numerous reports of success (Grichar and Boswell, 1987; Colvin and Brecke, 1988; Johnson et al., 2001; Tubbs and Gallaher, 2005). Additional research is needed to define factors that lead to inconsistent response to reduced tillage.
Another management technique likely to have interactive effects on peanut reproduction in either the ST or CT system is the irrigation application method used. Typical irrigation in peanut production is applied overhead with a lateral or pivot sprinkler system (OH); while another technique receiving some grower acceptance is the use of subsurface drip irrigation (SDI). Subsurface drip irrigation is purported to maximize water-use efficiency (WUE) by reducing soil evaporation, percolation, and runoff (Phene et al., 1992; Assouline et al., 2002). However, SDI systems, through the nature of their underground water application, may impede peg soil penetration due to drier soil surfaces and possibly elevated soil surface temperatures (Lanier et al., 2004; Sorensen et al., 2005). The presence of a cover crop in an ST system may ameliorate this problem and decrease soil surface evaporative losses while decreasing soil surface temperatures.
The objectives of this study were to: 1) quantify the differences in flower, peg, and pod formation between ST and CT systems; 2) determine if irrigation treatments within these tillage systems affected these same processes in both OH and SDI irrigation treatments; and 3) correlate climatic conditions with reproductive processes to determine their effects and interactions with tillage and irrigation treatments on peanut reproduction.
Material and Methods
Plot Design
The experiment was conducted during 2004 and 2005 at the Hooks Hanner Environmental Resource Center in Dawson, GA, US. Soil type at this site was a Greenville fine sandy loam (fine, kaolinitic, thermic Rhodic Kandiudults). Treatments included a factorial combination of two tillage systems (strip and conventional) and two irrigation methods (overhead and subsurface drip) arranged in a randomized complete block design. Tillage and irrigation treatments were completely randomized across the field and replicated three times for a total of 12 plots. Plots consisted of six 30 m rows spaced 91 cm apart, with the middle two rows used for yield evaluation. Peanut (cv Georgia Green) was planted on 6 May 2004 and 4 May 2005 in twin rows spaced 18 cm apart with an interplant distance of approximately 5 cm using a Monosem 8812 twin row vacuum planter (ATI Inc., Lexena, KS).
The conservation tillage system chosen was strip tillage (ST) which is the most commonly utilized form of conservation tillage in peanut production. Strip tillage (ST) plots were planted to rye (Secale cereale L. cv Wrens' Abruzzi) in the fall of 2003 and 2004. Rye was terminated with paraquat 30 d prior to peanut planting each year and followed by strip tillage. Strip tillage was performed with a single-pass implement (KMC Rip Strip, Kelley Mfg. Co., Tifton, GA) consisting of a coulter ahead of a subsoil shank, followed by two sets of fluted coulters ahead of a rolling basket and a drag chain assembly. An area of approximately 30 cm wide was tilled over the row. Conventional tillage (CT) plots remained fallow until the spring of each year at which time moldboard plowing to a depth of 25 cm was performed in both OH and SDI plots. Seedbed preparation followed with multiple diskings and/or field cultivations and the formation of a 91 cm raised bed. The irrigation treatments used in factorial combination with the tillage treatments included overhead (OH) and subsurface drip (SDI) irrigation. The OH treatment was applied using a lateral irrigation system that traveled across all of the twelve plots, but with water applied only to the randomized OH plots by blocking the sprinkler heads over the SDI plots. The SDI system consisted of drip tube laterals spaced 1.82 m apart between every other crop row. The drip tubing was 0.38 mm thick (15 mil) with emitters spaced at 45 cm and a flow rate per emitter of 0.77 l/h.
An on-site automated weather station (Hoogenboom, 1993) monitored and recorded environmental data including temperature, relative humidity, solar radiation, and wind speed and direction. Potential evapotranspiration (ETo) was calculated from the meteorological data using the Jensen-Haise equation (McGuinness and Bordne, 1972) corrected for local conditions. Daily plant water use (ETa) was calculated by muliplying ETp by a crop coefficient determined from DAP (Harrison and Tyson, 1993). Irrigation amount was calculated by subtracting the daily rainfall amount from the ETa. If rainfall was greater than ETa no irrigation was applied. Irrigation was scheduled daily for the SDI treatment, while irrigation amounts were summed over a 3–5 day period and then applied in the OH treatments. Therefore, the total water applied was the same in the SDI and OH treatments. Total irrigation applied in either the OH or SDI system was 282 mm in 2004 and 273 mm in 2005.
Plant Collection
Reproductive assessment was accomplished on two time scales in 2004 and 2005. Fine scale reproductive counts were conducted on a three to four day interval beginning 7 June 2004 and 8 June 2005 and ending 16 July 2004 and 8 July 2005, respectively. In 2004, three plants per plot were permanently tagged and the number of flowers was counted non-destructively. In addition, each flower was individually tagged using jewelry tags. The fate of each tagged flower was followed on a three day schedule and tagging of flowers was repeated at weekly intervals until 16 July 2004. Fine scale reproductive counts were conducted in 2005 by destructively sampling three plants per plot and counting the total number of flowers, pegs, and pods on the 3–4 day schedule. Therefore, for intensive counts performed every 3–4 days, flower number only was measured in 2004, while flowers, pegs, and pods were measured in 2005. Season-long counts were conducted in both 2004 and 2005 to quantify reproductive processes across the entire growing season. Number of flowers, pegs, and pods was determined on thirty day intervals starting 30 days after planting (DAP) and continuing until 120 DAP. At each sampling period, one plant per plot was destructively collected and reproductive structures counted.
Plots were harvested when the crop reached optimum maturity. Plots were mechanically inverted in 33 m by two row sections and machine threshed. Yield was determined after pods were dried to approximately 10% moisture. One kg subsamples were retained for farmer stock grade determination; pod yields are presented as weight per hectare minus foreign material.
Statistics
Data for reproductive counts were subjected to analysis of variance within 2004 and 2005 data separately to examine the effects of count date, tillage, irrigation, and all possible interactive effects on the difference in number of flowers, pegs, and pods. Significance levels were tested at p ≤ 0.05. Separate analyses were performed for the two data sets at different time scales: one analysis for the data collected every 3–4 d for a period of approximately six weeks, and one analysis for the data collected at 30 day intervals throughout the growing season. In 2004, flowers that were permanently tagged and monitored every 3–4 days for peg production, analysis of variance was performed on the percentage of flowers converted to pegs and the number of days it took to form a peg to determine the effects of tag date, tillage, irrigation, and all interactive effects.
Pearson correlations were analyzed between mean flower, peg and pod number per plant and weather parameters measured at the site. Weather conditions included: mean air temperature; mean relative humidity; soil temperature at 5, 10, and 20 cm depths; mean daily rainfall; incident radiation; and vapor pressure deficit. The same conditions taken 1, 3, and 6 days prior to reproductive counts were also analyzed to determine the effect of past edaphic conditions on reproductive processes. Correlations were run individually within year, tillage, and irrigation combinations.
Results
Fine Scale Counts of Flowers, Pegs, and Pods
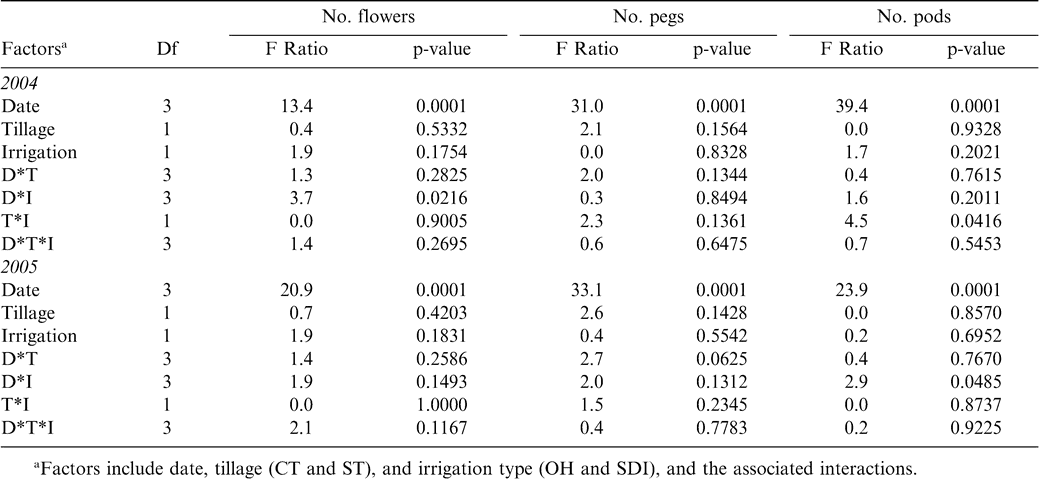
The period during which reproductive processes were monitored on a fine scale of 3–4 days captured both the initiation of flowers (Figure 1), pegs, and pods (Figure 2) as well as the eventual decline in flowering as evidenced by the significant effect of date during 2004 and 2005 for all traits measured (Table 1). Tillage and irrigation affected average flower number per plant in 2004 and the number of pods per plant in 2005. Average flower number per plant was higher in ST than in the CT treatment in 2004, while pod number per plant was greatest in OH irrigation than SDI in 2005. Significant interactions occurred between: date and tillage for 2004 flower counts; and date and irrigation, and tillage and irrigation for 2005 pod counts. The three-way interaction between date, tillage, and irrigation was only significant for 2005 flower counts (Table 1). While reproductive counts showed some differences between tillage and irrigation treatments, final plot yield did not show significant differences among treatments (Table 2).
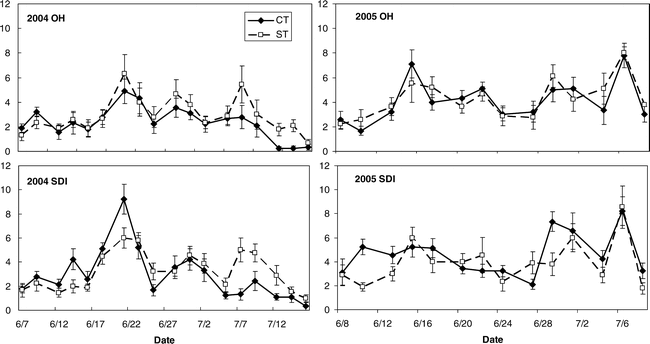
Mean number of flowers per plant in 2004 and 2005 for strip and conventional tillage (ST and CT) peanut under overhead and subsurface drip irrigation (OH and SDI).
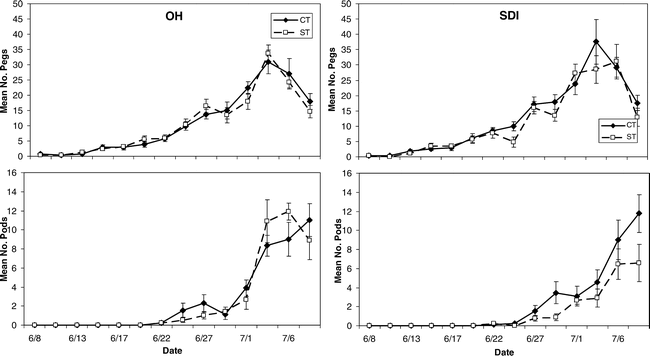
Mean number of pegs and pods in 2005 for strip and conventional tillage (ST and CT) peanut under overhead and subsurface drip irrigation (OH and SDI).
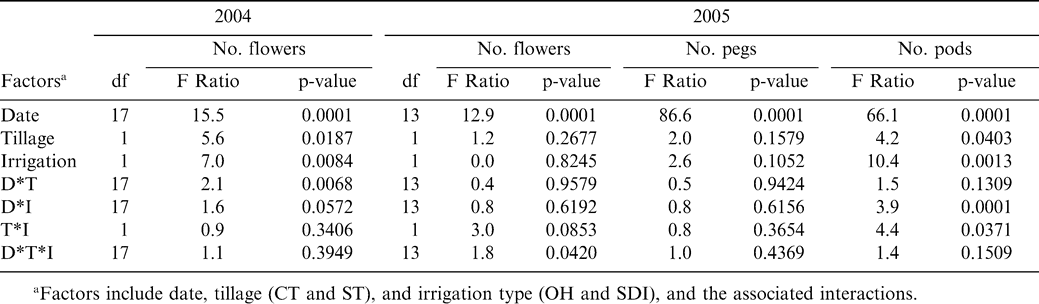
Analysis of variance for average number of flowers (2004 and 2005), pegs, and pods (2005) counted every 3–4 days.

Plot yields for 2004 and 2005 seasons. Numbers are reported for both conventional and strip tillage plots under overhead (OH) and subsurface drip (SDI) irrigation.
Peak flowering time period in 2004 for both tillage and irrigation treatments was between 19 and 23 June and diminished later in the season (Figure 1). In contrast, flowering appeared to peak nearly a month later in 2005 (6 July). The OH irrigation treatment also showed an earlier flowering peak for both CT and ST (near 14 June 2005) than for 2004 when the peak was near 22 June (Figure 1). Because flowers are anatomical precursors to pegs and pods, average counts for these structures necessarily peaked much later than flower counts during the monitoring time period in 2005. Maximum peg counts in the OH and SDI irrigation treatments in both tillage treatments occurred between 4 July and 6 July 2005 (Figure 2). Pod number per plant in the OH treatment under ST tillage peaked directly after the pegs did on approximately 6 July; while in the OH-CT, SDI-CT, and SDI-ST treatments, pod number per plant did not decline up to the last monitoring date (Figure 2).
Season-long Counts
While the intensive reproductive counts were able to capture the detailed rise and fall of flower production, season-long counts showed the full production curve for flowers, pegs, and pods (Figure 3). No differences in reproductive counts were noted among tillage or irrigation in either 2004 or 2005 (Table 3). Collection date was significant for all tissue types and for both years as expected due to the effect of seasonal variation in these characteristics. Significant interactions in 2004 included: date by irrigation for flowers; and tillage by irrigation for pods. In 2005, pods had a significant interaction between date and irrigation. The significant effect of date on seasonal counts was illustrated by differences in peak production over the season (Figure 3). In both the OH and SDI treatment, flower production showed a main peak in July 2004 and 2005 for both tillage treatments, with a second flush of flowers occurring in September 2004. The OH treatment showed a peak in peg production in the ST treatment in August 2004; while the ST and CT treatments peaked in July 2005 with the CT counts greater in magnitude than the ST treatment. The SDI irrigation treatment had similar maximum peg count patterns but with somewhat lower peg numbers. Pod counts showed steady linear patterns with the first pods evident on the July collection. Pod counts for all years, irrigation, and tillage treatments continued to rise through September in both years (Figure 3).
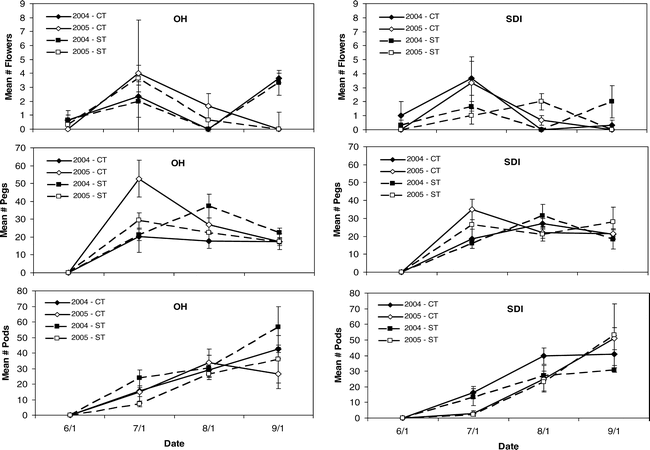
Seasonal reproductive counts conducted throughout the growing season for mean number of flowers, pegs, and pods per plant in strip and conventional tillage (ST and CT) under overhead and subsurface drip irrigation (OH and SDI). Counts were conducted at approximately 30, 60, 90, and 120 days after planting.
Analysis of variance for average number of flowers, pegs, and pods counted over the entire growing season on a 30 day basis in 2004 and 2005.
Correlations with Weather Conditions
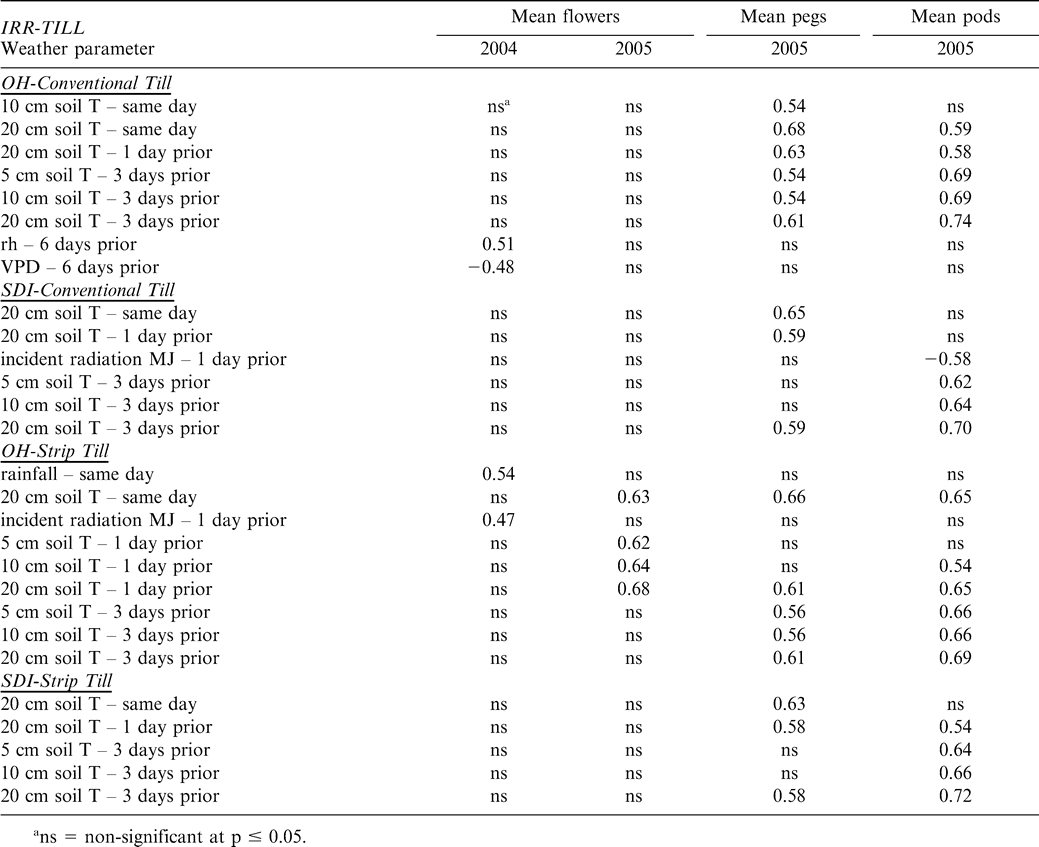
Across all tillage and irrigation treatments in 2004 and 2005, very few weather conditions were correlated to flower production; only the OH-CT and the OH-ST treatments showed significant correlations between weather conditions and flowering, while both SDI treatments showed no significant correlations (Table 4). For 2004, in the OH-CT treatment, relative humidity and vapor pressure deficit six days prior to flower counts were correlated, while in the OH-ST treatment, rainfall measured the same day and incident radiation measured one day prior to flower counts were correlated. In 2005, soil temperature (both same day and one day prior to flower counts) was highly correlated to flower counts but only in the OH-ST treatment. Soil temperature was the primary weather factor affecting peg and pod production. For pegs, soil temperature at 5 cm, 10 cm, and 20 cm had effects in all tillage and irrigation combinations (Table 4). Effects were important the same day, one and three days prior to reproductive counts. Typically, weather conditions one to three days prior to counts were well correlated with pod count per plant (Table 4).
Pearson correlations between weather parameters and mean flowers, pegs, and pods per plant. Weather parameters were measured the same day and 1, 3, and 6 days prior to reproductive counts. Significant correlations are reported for each year (2004, 2005), tillage (conventional and strip), and irrigation (OH, SDI) combination. Pegs and pods were not counted in 2004.
Tagged Flowers
When flowers were tagged and monitored over time in 2004, differences in the percent of flowers that produced a peg and the number of days it took to form those pegs showed very limited differences among treatments. Tillage affected the percent of flowers that were converted to pegs with the CT treatment having a higher percentage conversion than ST (df = 1, F Ratio = 6.5, p-value = 0.0156). In addition for this trait, the interactive effects of tillage by irrigation (df = 1, F Ratio = 6.4, p-value = 0.0166) showed significant effects. The percent of tagged flowers converted to pegs in 2004 showed no large numerical differences between tillage or irrigation treatments until the last monitoring date on 2 July (Figure 4). Overall, the numerical percentages showed a lower percentage of OH-ST flowers were converted to pegs followed by SDI-CT, SDI-ST, and OH-CT with the highest percent conversion. Weather parameters affecting percent peg conversion occurred almost exclusively six days prior to the monitoring time period (Table 5). For the OH-ST treatment, relative humidity, incident radiation, and vapor pressure deficit six days prior to reproductive counts affected the conversion of flowers to pegs; while the SDI-CT treatment showed only relative humidity six days prior was important. Only the OH-CT treatment showed a same day response of rainfall in a negative direction with percent peg conversion (Table 5). The number of days it took for a flower to produce a peg showed a significant effect of date on which the flower was tagged (df = 3, F Ratio = 4.1, p-value = 0.0077) and the interaction between tillage and irrigation (df = 1, F Ratio = 5.7, p-value = 0.0179). The interaction between tillage and irrigation was due to the longer time for flower to peg conversion in the ST plants under SDI irrigation, while ST plants under OH irrigation exhibited a shorter conversion time period (Figure 5).
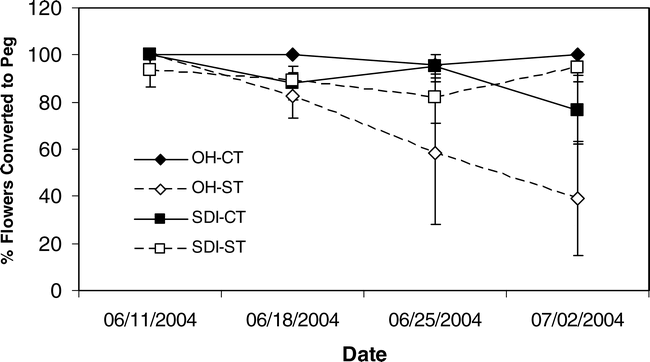
Percent of flowers converted to pegs in 2004 for the combination of overhead and subsurface drip irrigation (OH and SDI) with strip and conventional tillage (ST and CT) treatments. “Date” is the day when the flowers were tagged.
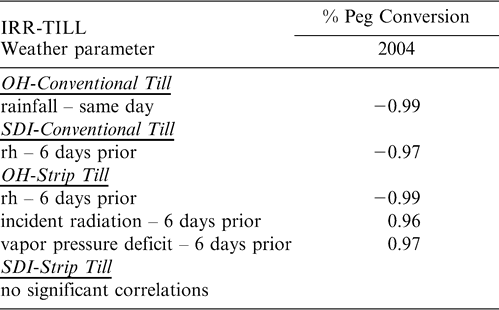
Pearson correlations between weather parameters and percent of flowers converted to pegs during the measurement period in 2004. Weather parameters were measured the same day and 1, 3, and 6 days prior to reproductive counts. Significant correlations are reported for each tillage (conventional and strip), and irrigation (OH, SDI) combination.
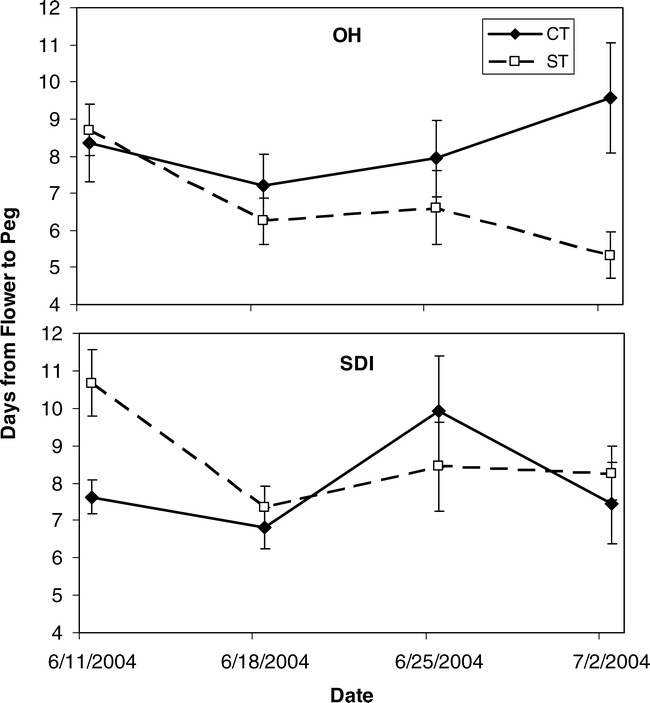
Mean number of days from flower appearance to peg formation in 2004 for strip and conventional tillage (ST and CT) peanut under overhead and subsurface drip irrigation (OH and SDI).
Discussion
This study has provided a unique examination of the effects of both tillage and irrigation management systems on peanut reproduction. Overall, the study finds minimal impact of tillage systems on peanut reproduction and no resultant decrease in yield associated with conservation tillage over the two year study period. Due to the increasing number of producers either considering or currently converting conventional tillage practices into conservation tillage, this knowledge can aid growers in this decision. While conservation tillage has shown some variability in its effect on peanut yield and quality among previous studies, the current results add evidence that peanut reproduction appears not to be adversely affected in ST systems. Another inimitable opportunity this study has provided is the comparison of results from previous studies of the environmental effects on peanut reproduction, conducted almost exclusively in greenhouse environments, with actual quantification of these processes in a typical field peanut production environment. The results show that the reproductive processes occurring in the artificial environment vs. the production environment are not always comparable.
Both fine scale and season-long measurements of flowers, pegs, and pods show very little difference when comparing growth in ST and CT systems. Despite grower fears, flower production at a 3–4 day time scale in the ST system in 2004 was greater than the CT system. However, seasonal counts showed no differences in flower, peg, or pod production between the two systems in either year. This was likely due to the lower percentage of flowers converted to pegs in the ST system, thereby leading to no real differences between the two tillage treatments in peg or pod numbers. In both tillage systems, the average time from flower appearance to peg production was within the typical range of 6–9 days (Vara Prasad et al., 2001). The only other difference seen between the CT and ST systems was for pod production in 2005 where CT plants produced, on average, 2.5 pods per plant in comparison to 2.1 pods per plant for those grown under and ST system. Despite these differences in pod production over a two week period, season long pod counts and eventual total pod production (yield) showed no differences between the two tillage systems.
The effects of irrigation in the current study showed that the choice of application method was important to the maintenance of reproductive output. While the SDI treatment showed increased numbers of flowers in 2004, it showed fewer numbers of pods than the traditional OH irrigation treatment in 2005. But the true disadvantage to growing peanuts under subsurface drip irrigation appeared to be when it was utilized in combination with conservation tillage. The significant interaction between tillage and irrigation method showed, on average, the SDI-CT treatment produced 2.4 pods per plant in comparison to 1.5 pods per plant in the SDI-ST treatment. This interactive effect was robust enough to be seen in the season-long counts as well, such that the SDI-ST treatment had depressed pod production in comparison to the SDI-CT treatment. While yields were not lowered in the SDI-ST treatment in the two years of the study, conservation tillage may put peanuts produced in this system at higher risk for decreased pod production.
By necessity, almost all of the experiments seeking to determine the fine scale effects of air temperature, soil temperature, and other environmental factors on reproductive processes in peanut have been conducted in controlled environmental chambers - either greenhouses or polyethylene covered plots. While these studies have discovered details about the formation of flowers, pegs, and pods, their utility in actual production environments may be limited due to the vast differences between the greenhouse environment and the field climate, and how these two different environments affect phenotypic plasticity in peanuts. The current study certainly cannot make conclusions about all possible environmental conditions experienced in a field setting, but it does provide a timely and much needed opportunity to examine the correlation between climatic conditions and peanut reproduction under field conditions and apply this information to grower management decisions. Surprisingly, the results show limited direct effects of atmospheric conditions on flower, peg, and pod production. This is somewhat contrary to what has been found previously in greenhouse environments, where high air temperatures can increase the total number of flowers (Vara Prasad et al., 1999b). However, ambient air conditions can affect more than just flower number through changes in flower morphology itself including: increasing the length of the hypanthium in the flower; increasing stigma exertion and reducing the rate of successful germination; reducing the rate of pollen tube growth; and decreasing the number of pegs produced (Ketring, 1984; Talwar et al., 1999; Talwar and Yanagihara, 1999). The primary effect of atmospheric conditions in this study was on peg production; in this case, conditions six days prior to peg appearance had the greatest effect. These conditions, including relative humidity and vapor pressure deficit (Table 5) were likely affecting microsporogenesis which occurs three to six days before anthesis and may be highly sensitive to temperature (Xi, 1991; Vara Prasad et al., 1999b). Therefore, heat stress is likely to affect the conversion rate of flowers into pegs, but may not affect actual flower production or the conversion of pegs into pods (Vara Prasad et al., 1999b). Optimum air temperatures for vegetative growth and physiological performance of peanut is in the range of 25–35 C but for reproductive processes is in the range of 20–25 C (Williams and Boote, 1995; Talwar et al., 1999; Talwar and Yanagihara, 1999). However, a threshold of 34 C has been demonstrated to exist, beyond which most of the deleterious effects on pollen production and viability occur (Vara Prasad et al., 1999a; Kakani et al., 2002). An examination of the atmospheric conditions during the current study indicates the absence of extreme heat stress in either 2004 or 2005 (Figure 6), thereby making negative effects of climate on flowers and pollen unlikely.
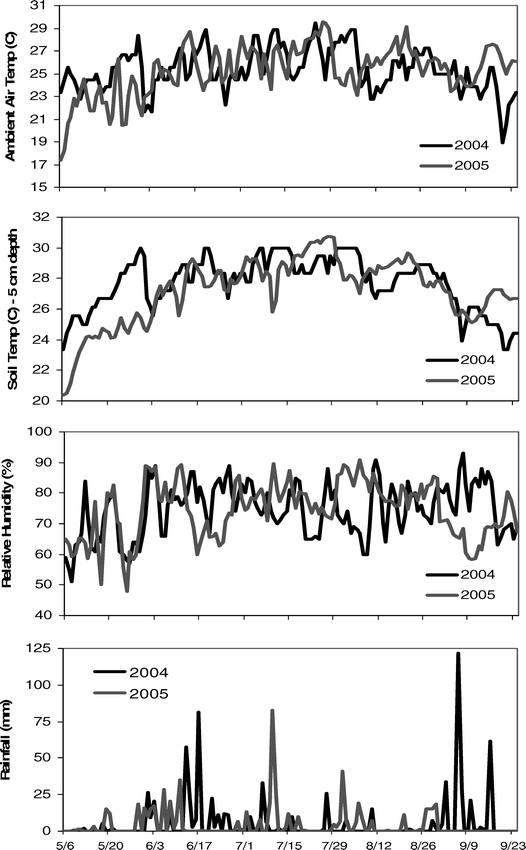
Average weather patterns for the 2004 and 2005 growing seasons.
What was clear in the current results was the effect of soil temperature on peanut reproduction, both the current day conditions and 1–6 days prior to reproductive counts. Environmental effects also appeared to act equally among tillage and irrigation treatments. Both peg and pod counts were significantly correlated with soil temperatures in the pod zone (5 cm) and below (10, and 20 cm) in all of the irrigation-tillage treatments. Further, all correlations were such that reproductive production increased with increasing soil temperature. High soil temperatures have been found to significantly shorten times to flowering, pod formation, and maturity, but to decrease total pod number, pod weight, and total yield as temperatures range from 23–35 C (Dreyer et al. 1981; Golombek et al., 2001; Awal et al., 2003; Craufurd et al., 2003). The reason may be that pods at high temperatures may not be able to accept assimilates very quickly and the rate of translocation of photosynthates may be slowed (Awal et al., 2003). However, one study found a positive correlation between pod weight and temperature in the range of 10–30 C (Ono et al., 1974), so there appears to be an optimal intermediate temperature regime where pod development and soil temperature are positively correlated. When temperatures either are below or above this point, pod production decreases (Golombek and Johansen, 1997). Looking at the soil temperature in this study at the pod depth (5 cm), temperatures fell exclusively within the optimal range making positive correlations with pod production likely (Figure 6). Interestingly, significant differences between 2004 and 2005 in plot yield can possibly be explained by the large variation in soil temperatures between the two years during early establishment and primary reproductive activity (6 May – 10 June). Soil temperatures during this time period were much higher in 2004 than 2005, leading to better pod development and production in the former year.
Based on the results in this study, conservation tillage in peanuts appears to be a viable production strategy. Previous research has already demonstrated that conservation tillage systems can spell great benefits to producers through decreased soil losses, increased soil water infiltration, and increased plant available water. In this study, no deleterious effects of conservation tillage on peanut reproduction were noted except in the subsurface drip irrigation treatment. Therefore, for the majority of U.S. peanut growers that utilize overhead irrigation, conservation tillage remains a beneficial alternative to conventional clean till production.
Acknowledgements
We thank Kathy Gray, Ashley Butts, William Pearce, and Latoya Rucker for their meticulous field assistance. We thank the Hooks Hanner Environmental Resource Center and the Georgia Soil and Water Conservation Commission for the field space and D. Eigenberg for research support.
Literature Cited
Assouline S. , Cohen S. , Meerback D. , Harodi T. , and Rosner M. 2002 Microdrip irrigation of field crops: effect on yield, water uptake, and drainage in sweet corn. Soil Sci. Soc. Am. J 66 : 228 – 235 .
Awal M. A. , Ikeda T. , and Itoh R. 2003 The effect of soil temperature on source-sink economy in peanut (Arachis hypogaea). Environmental and Experimental Botany 50 : 41 – 50 .
Blevins R. L. , Cook D. , Phillips S. H. , and Phillips R. E. 1971 Influence of no-tillage on soil moisture. Agronomy J 63 : 593 – 596 .
Cantonwine E. G. , Culbreath A. K. , Stevenson K. L. , Kemerait R. C. , Brenneman T. B. , Smith N. B. , and Mullinix B. G. 2006 Integrated Disease Management of Leaf Spot and Spotted Wilt of Peanut. Plant Disease 90 : 493 – 500 .
Colvin D. L. and Brecke B. J. 1988 Peanut cultivar response to tillage systems. Peanut Sci 15 : 21 – 24 .
Craufurd P. Q. , Prasad P. V. V. , Kakani V. G. , Wheeler T. R. , and Nigam S. N. 2003 Heat tolerance in groundnut. Field Crops Res 80 : 63 – 77 .
CTIC (Conservation Technology Information Center) 2006 2006 Crop Residue Management Survey. Available online only: http://www.conservationinformation.org/pdf/2006CRMSurveySummaryHiRes.pdf.
Dreyer J. , Duncan W. G. , and McCloud D. E. 1981 Fruit temperature, growth rates, and yield of peanuts. Crop Sci 21 : 686 – 688 .
Gallaher R. N. 1977 Soil moisture conservation and yield of crops no-till planted in rye. Soil Sci. Soc. Am. J 41 : 145 – 147 .
Golombek S. D. and Johansen C. 1997 Effect of soil temperature on vegetative and reproductive growth and development in three Spanish genotypes of peanut (Arachis hypogaea L.). Peanut Sci 24 : 67 – 72 .
Golombek S. D. , Sultana A. , and Johansen C. 2001 Effect of separate pod and root zone temperatures on yield and seed composition of three Spanish cultivars of groundnut (Arachis hypogaea L.). J. Sci. Food Agric 81 : 1326 – 1333 .
Grichar W. J. and Boswell T. E. 1987 Comparison of no-tillage, minimum, and full tillage cultural practices on peanuts. Peanut Sci 14 : 101 – 103 .
Harrison K. and Tyson A. 1993 Irrigation Scheduling Methods. Bulletin – Cooperative Extension Service, Univ. of GA, College of Agriculture, 12 p.
Hoogenboom G. 1993 The Georgia Automated Environmental Monitoring Network, Southeastern Climate Review 4 / 1 : 12 – 18 .
Johnson W. C. , Brenneman T. B. , Baker S. H. , Johnson A. W. , Sumner D. R. , and Mullinix B. G. 2001 Tillage and pest management considerations in a peanut-cotton rotation in the southeastern coastal plain. Agronomy J 93 : 570 – 576 .
Jordan D. L. , Barnes J. S. , Bogle C. R. , Brandenburg R. L. , Bailey J. E. , Johnson P. D. , and Culpepper A. S. 2003 Peanut response to cultivar selection, digging date, and tillage intensity. Agron. J 95 : 380 – 385 .
Kakani V. G. , Prasad P. V. V. , Craufurd P. Q. , and Wheeler T. R. 2002 Response of in vitro pollen germination and pollen tube growth of groundnut (Arachis hypogaea L.) genotypes to temperature. Plant, Cell and Envrironment 25 : 1651 – 1661 .
Ketring D. L. 1984 Temperature effects on vegetative and reproductive development of peanut. Crop Sci 24 : 877 – 882 .
Lanier J. E. , Jordan D. L. , Barnes J. S. , Mattthews J. , Grabow G. L. , Griffin W. J. , Bailey J. E. , Johnson P. D. , Spears J. F. , and Wells R. 2004 Disease management in overhead sprinkler and subsurface drip irrigation systems for peanut. Agronomy J 96 : 1058 – 1065 .
Marois J. J. and Wright D. L. 2003 Effect of tillage system, phorate, and cultivar on tomato spotted wilt of peanut. Agronomy J 95 : 386 – 389 .
McGuinness J. L. and Bordne E. F. 1972 A comparison of lysimeter-derived potential evapotranspiration with computed values. U.S. Department of Agriculture, Agricultural Research Service Technical Bulletin 1452
Merrill S. D. , Black A. L. , Fryrear D. W. , Saleh A. , Zobek T. M. , Halvorson A. D. , and Tanaka D. L. 1999 Soil wind erosion hazard of spring wheat-fallow as affected by long-term climate and tillage. Soil Sci. Soc. Am. J 63 : 1768 – 1777 .
Monfort W. S. , Culbreath A. K. , Stevenson K. L. , Brenneman T. B. , Gorbet D. W. , and Phatak S. C. 2004 Effects of reduced tillage, resistant cultivars, and reduced fungicide inputs on progress of early leaf spot of peanut (Arachis hypogaea). Plant Disease 88 : 858 – 864 .
Ono Y. , Nakayama K. , and Kubota M. 1974 Effects of soil temperature and soil moisture in podding zone on pod development of peanut plants. Proc. Crop Sci. Soc. Japan 43 : 247 – 251 .
Phene C. J. , Hutmacher R. B. , El Ayars J. , Davis K. R. , Mead R. M. , and Schoneman R. A. 1992 Maximizing water use efficiency with subsurface drip irrigation. ASAE Paper No. 92-2090 St. Joseph, Mich ASAE .
Phillips R. E. , Blevins R. L. , Thomas G. W. , Frye W. W. , and Phillips S. H. 1980 No-tillage agriculture. Science 208 : 1108 – 1113 .
Raper R. L. , Reeves D. W. , Schwab E. B. , and Burmester C. H. 2000 Reducing soil compaction of Tennessee Valley soils in conservation tillage systems. J. Cotton Sci 4 : 84 – 90 .
Reeves D. W. 1997 The role of soil organic matter in maintaining soil quality in continuous cropping systems. Soil and Tillage Res 43 : 131 – 167 .
Sorensen R. B. , Butts C. L. , and Rowland D. L. 2005 Five years of subsurface drip irrigation on peanut: what have we learned? Peanut Sci 32 : 14 – 19 .
Talwar H. S. , Takeda H. , Yashima S. , and Senboku T. 1999 Growth and photosynthetic responses of groundnut genotypes to high temperature. Crop Sci 39 : 460 – 466 .
Talwar H. S. and Yanagihara S. 1999 Physiological basis of heat tolerance during flowering and pod setting stages in groundnut (Arachis hypogaea L.). JIRCAS Working Report No 14 : 47 – 65 .
Tubbs R. S. and Gallaher R. N. 2005 Conservation tillage and herbicide management for two peanut cultivars. Agronomy J 97 : 500 – 504 .
Vara Prasad P. V. , Caufurd P. Q. , Kakani V. G. , Wheeler T. R. , and Boote K. J. 2001 Influence of high temperature during pre- and post-anthesis stages of floral development on fruit-set and pollen germination in peanut. Aust. J. Plant Physiol 28 : 233 – 240 .
Vara Prasad P. V. , Craufurd P. Q. , and Summerfield R. J. 1999a Fruit number in relation to pollen production and viability in groundnut exposed to short episodes of heat stress. Annals of Botany 84 : 381 – 386 .
Vara Prasad P. V. , Craufurd P. Q. , and Summerfield R. J. 1999b Sensitivity of peanut to timing of heat stress during reproductive development. Crop Sci 39 : 1352 – 1357 .
Williams J. H. and Boote K. J. 1995 Physiology and modeling – predicting the unpredictable legume. 301 – 353 In Pattee H. E. and Stalker H. T. Advances in Peanut Science Stillwater, OK, USA American Peanut Research and Education Society .
Xi X. Y. 1991 Development and structure of pollen and embryosac in peanut (Arachis hypogaea L.). Bot. Gaz. (Chicago) 152 : 164 – 172 .
Yu B. , Sombatpanit S. , Rose C. W. , Ciesiolka C. A. A. , and Coughlan K. J. 2000 Characteristics and modeling of runoff hydrographs for different tillage treatments. Soil Sci. Soc. Am. J 64 : 1763 – 1770 .
Notes
- Plant Physiologist, Research Agronomist, and Agricultural Engineer, USDA-ARS, National Peanut Research Laboratory, PO Box 509, 1011 Forrester Dr. SE, Dawson, GA 39842 [^]
- Corresponding author (email: drowland@nprl.usda.gov). [^]
Author Affiliations
Disclaimer
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA.